
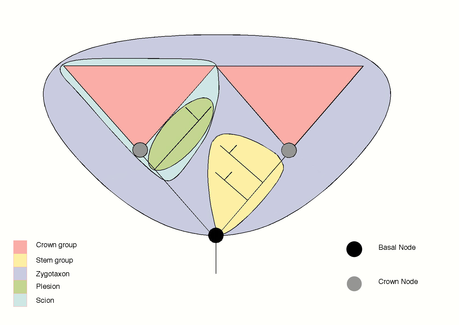
クラウングループ(英:crown group) とは、系統学において、ある系統で現生する種の最も近い共通祖先の子孫全てから構成される系統群のこと。
この概念は、 系統学や分岐学の創始者であるヴィリー・ヘニッヒにより、現生種を絶滅した近縁生物につなげて分類する方法として、彼の『昆虫の系統発生』(独:Die Stammesgeschichte der Insekten)の中で展開され、「クラウン(冠)」や 「ステム(幹)」という系統群を表す用語は1979年にRPS Jefferiesによって作られた 。
1970年代に確立されたが、2000年代にグレイアム・バッドらによって再度紹介されるまで、一般に認知されることはなかった。
概要
系統樹において現生種の最も近い共通祖先から派生する部分には、現生種のみからなる枝も、絶滅種のみからなる枝も存在するが、いずれもクラウングループの一部である。例えば、仮にクラウン鳥類を考える(すなわち、全ての現生鳥類の最も近い共通祖先の子孫を現生・絶滅問わず考える)と、ドードーやオオウミガラスのような絶滅種からなる枝も、現生する全鳥類の最近共通祖先から伸びたもので、鳥類のクラウングループの中に収まる。鳥類についての非常に単純化された系統樹を以下に示す。
| 鳥綱 |
|
||||||||||||
この系統樹において 「新鳥類」とされる系統群が鳥類のクラウングループであり、全ての現生する鳥類の最も近い共通祖先とその子孫が現生・絶滅問わず含まれる。鳥綱の中でも始祖鳥や「他の絶滅した群」とされている部分は鳥類のクラウングループに含まれない。
現生種を使わずに、問題のクラドゲネシス(系統分化)から生じたと定義することもできる。
クラウングループで分類群を考える時は、一般に定義されている分類群と区別するために、「クラウン〇〇」というように表現されることが多い。例えば、鳥類と哺乳類の伝統的な定義はそれらの特徴によるもので、現生種の最も新しい共通祖先以前の絶滅種や、哺乳動物ハルダノドンなど、後年に生息してはいるが問題の共通祖先から派生していない絶滅種も含んでいる。このため、クラウン鳥綱とクラウン哺乳綱は、伝統的な定義の鳥綱と哺乳綱とは内容が若干異なり、文献で若干の混乱を生んでいる。
クラウングループの概念下にある別のグループ
分岐学では、グループの定義は系統樹のトポロジー(系統関係、分岐順序)に厳密に従うが、特に化石種を考える場合、現生生物を用いて系統樹の一部を明確に表現するため、クラウン以外のいくつかの接頭語が定義されている。
パングループ
パングループ(pan-group)またはトータルグループ(total group)は、クラウングループに加え、クラウングループ外の現生生物よりもクラウングループに近縁な全ての絶滅種を含めたものである。系統樹では、クラウングループとその次に近い現生種とを繋ぐ節から、クラウングループ側に伸びる枝全てとなる。
パングループとしての鳥綱には、新鳥類に加え、ワニ目よりも鳥類に近縁関係にある全ての生物が含まれる。新鳥類からワニ目の系統と合流する地点まで戻る系統樹が、全ての側枝も共に含めてパングループとしての鳥綱を構成する。ここには始祖鳥やヘスペロルニスや孔子鳥のようにクラウングループ外の原始的な鳥に加え、恐竜や翼竜、マラスクスなども含まれる。
パングループとしての哺乳綱は、全ての哺乳類および残りの有羊膜類(竜弓類)との分岐以降の哺乳類に近縁な生物で構成され、単弓類と同義になる。
ステムグループ
ステムグループ(stem group)はパングループから、クラウングループ自体を除いた(つまり全ての現生種を除く)もので、必然的に側系統群である。これは、クラウングループの最も新しい共通祖先からそれらに最も近い現生の近縁種の間にある、クラウングループに近縁な原始的な生物からなると言える。定義上、ステムグループは絶滅種のみで構成される。 「ステムグループ」はクラウングループに関連した概念の中では最も頻繁かつ使用され、かつ最も重要なものである。化石種はクラウングループに含まれないため、これらを系統に沿って分類したり議論したりする際にはステムグループを使うことになる。 (上の系統樹で、黄色で表示された箇所がステムグループ。)
ステムグループの概念はジェフリーズ(1979)が初出とされることもあるが、ウィルマン(2003)によれば実際はドイツの体系学者オスニール・アベル(1914) が初出であり、1933年にアルフレッド・ローマーにより英語で早期に議論され、図示までされているという。
また、「ステムグループ」はクラウングループ外の中でも伝統的にその分類群に含まれるものだけを指す、狭い意味で用いられることもある。この場合、ステム哺乳綱にディメトロドンやアンテオサウルスのようなペルム紀の単弓類は含まれないことになる。
ステムグループの例
ステム鳥類(Stem birds)は、系統関係がかなりよく知られていることもあり、ステムグループとしてはおそらく最も用例が多い。以下の系統樹はベントン(2005)に基づく。
| 主竜類 |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||
※翼竜から始祖鳥までのグループが「鳥類のステムグループ」。古顎類と新顎類のグループが「鳥類のクラウングループ」。
ここでのクラウングループは新鳥類であり、現代のあらゆる鳥の系統は彼らの最も新しい共通祖先に遡る。鳥の最も近い現生生物はワニ目である。系統樹を新鳥類から左にたどると、ワニ目と合流するまでの全ての枝がステム鳥類に属する。ここには始祖鳥、ヘスペロルニス、孔子鳥のようなクラウングループ外の原始鳥類に加えて、恐竜と翼竜が含まれる。鳥とワニ目の最も新しい共通祖先(ここから派生するクラウングループは主竜類に相当する)は、鳥とワニいずれの特徴も有していなかった。鳥類のステムグループが進化するにつれて、羽や含気骨(または中空骨)といった鳥ならではの特徴が現れた。ステムグループの末端部からクラウングループの基部に移行した段階では、現生する鳥類に共通する全ての特徴が存在していた。
ステム哺乳綱(Stem mammals)は現生する哺乳類につながる系統で、竜弓類と分岐してから現生哺乳類の最も新しい共通祖先に至るまでの系統と、そこから分岐する枝をまとめたものとなる。このグループには単弓類に加え、モルガヌコドン目や梁歯目のような哺乳形類が含まれている。哺乳形類は哺乳類のクラウングループ外ではあるが、伝統的には解剖学的な観点から哺乳類とみなされている。
ステム四肢動物(Stem Tetrapoda)は、魚類側で最も近縁な種である肺魚と分岐してから四肢動物につながる系統に属する動物である。ここには肉鰭綱の一部に加えて、初期の迷歯亜綱の一部も含まれる。初期の四肢動物の系統関係は十分に分かっていないため、四肢動物のステムグループ内の迷歯亜綱にどの生物が含まれるのかは、正確には不明瞭である。これは、系統に関する定説が得られていない場合、クラウングループとステムグループを定義することの意義が限定的になることを表す一例である。
ステム節足動物(Stem arthropods)は、カンブリア紀のバージェス動物群とシリウスパセット動物群で見つかる古生物をはじめとして注目されているグループである。パンブデルリオン、ケリグマケラ、オパビニアとラディオドンタ類(アノマロカリス、フルディアなど)は、節足動物の特徴の一部を有しており、ステム節足動物と考えられている。これらの古生物を節足動物のステムグループに分類することで系統的な議論が可能になり、例えば節足動物の複眼と先頭の外骨格は胴部の外骨格より早期に起源することや、節足動物は緩歩動物(クマムシ)や有爪動物(カギムシ)と同様に葉足動物から派生するという系統仮説がこれで有力視されるようになった。
ステム鰓曳動物(Stem priapulids)は、初期カンブリア紀から中期のカンブリア紀の他の動物群で、澄江動物群やバージェス動物群で見つかっている。オットイアは現代の鰓曳動物とほぼ同じ構造をしているが、系統解析ではクラウングループ外となり、ステム鰓曳動物と考えられる。
プレシオングループ
プレシオン(plesion、ギリシャ語で「近い」の意)という言葉は系統分類学では長年にわたって使われたものであるため、プレシオングループは多義語となっている。 例としては「近隣のグループ」、すなわち、ある分類群の姉妹群を、クラウングループかどうかにかかわらず指すというものがある。この用語はまた、原始的特徴(共有原始形質)により定義される、側系統群を意味することもある。一般的には、問題のグループよりも早く分岐した枝を指す(最初の図の緑色)。
古生物学的な意義
化石生物を正しくステムグループに属させることで、それらの関係は(属させた研究者等の考えに基づくものとして)明確に示されることになり、問題のグループにおける主要な特徴の進化がどのような生態学・機能的な背景を持っていたかも明らかになる。そのため、ステムグループは古生物学の情報と現生生物の進化を統合して考える上で重要である。また、現生グループの特徴を示さないために別の独立したグループにあると考えられていた化石群も、ステムグループの一員と認識することで、現生生物との関連が明示される。そのような化石は、四肢動物、哺乳動物、動物などの起源を考える上で特に重要であった。
ステムグループの概念の適用は、バージェス動物群の解釈にも影響を与えた。独立した門ではなく、現存する門のステムグループに分類することで、カンブリア爆発をより理解しやすくなり、特殊な進化メカニズムを持ち出す必要がなくなったとも考えられているが、カンブリア爆発の唐突さを解明しようとする理論進化学者の困難が改善されたわけではない。また、ステムグループという概念を過剰に強調することで、新たな高等分類群の適切な認識を遅らせたり、不明瞭にする恐れもある。
系統分類学におけるステムグループ
カール=エルンスト・ローターバッハによって提案されたように、ステムグループには接頭語「ステム」(すなわち、ステム鳥綱、ステム節足動物)を付すべきだが、クラウングループには接頭語をつけるべきではない。この考えは必ずしも全ての研究者の間で共有されてはいないものの、多くの古生物学者はこれに倣っている。
脚注
注釈
関連文献
- Budd, G. (2001), “Climbing Life's Tree”, Nature 412 (6846): 487, doi:10.1038/35087679, PMID 11484029
- Budd, G. E. (2001), “Tardigrades as 'stem-group' Arthropods: the Evidence from the Cambrian Fauna”, Zoologischer Anzeiger 240 (3?4): 265?279, doi:10.1078/0044-5231-00034
- Craske, A. J.; Jefferies, R. P. S. (1989), “A New Mitrate from the Upper Ordovician of Norway, and a New Approach to Subdividing a Plesion”, Palaeontology 32: 69?99