
| ツノゴケ植物門 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ニワツノゴケ属の一種
(Phaeoceros carolinianus) | |||||||||||||||
| 分類 | |||||||||||||||
| |||||||||||||||
| 学名 | |||||||||||||||
|
Anthocerotophyta Stotler & Stotl.-Crand. | |||||||||||||||
| 英名 | |||||||||||||||
| hornworts, anthocerotes | |||||||||||||||
| 下位分類 | |||||||||||||||
ツノゴケ類(ツノゴケるい) (英:hornworts) は陸上植物の一群であり、蘚類や苔類とともにコケ植物とよばれる。本体である配偶体は茎や葉を欠く扁平な葉状体であり、そこから細長い角状の胞子体が生じる。和名の「ツノゴケ (角苔)」や英名の「hornworts」は、いずれもこの角状の胞子体の形に由来する。各細胞がふつうピレノイドを含む葉緑体を1個だけもつという点で、陸上植物の中で特異な存在である。2019年現在、分類学的にはふつうツノゴケ植物門 (学名: Anthocerotophyta) に分類される。およそ150種ほどが知られ、日本からは6属17種が報告されている。
特徴
配偶体
他のコケ植物と同様、ツノゴケ類の生活環の主な世代は配偶体 (ゲノムを1セットだけもち配偶子を形成する体) である。ツノゴケ類の配偶体は茎や葉をもたず、扁平なロゼット状またはリボン状の葉状体である (下図)。ツノゴケ類の葉状体はゼニゴケ (苔類) の葉状体のような複雑な組織分化を示さない比較的単純な構造をしており (単純葉状性)、苔類の中ではミズゼニゴケなどに似ている。種によっては、葉状体に中肋をもつ (例:キノボリツノゴケ属)。葉状体は基本的に多細胞層であるが、縁辺部 (翼部) が単細胞層である種もいる。


葉状体の腹面からは、多数の仮根が生じ、基物に付着している。仮根は単細胞性であるが、先端が分岐することがある。また腹面の中軸沿いにはふつう孔 (slime pore, mucilage cleft) がある。この孔は気孔のように2個の細胞で囲まれているが、気孔との相同性は明らかではない。
葉状体内には、粘液質で満たされた細胞間隙 (mucilage canal, mucilage chamber) が存在する。この間隙の中には、しばしば窒素固定能をもつ藍藻 (ネンジュモ属) が細胞外共生している。藍藻は葉状体腹面の孔 (上記) から侵入し、間隙内で増殖する。スジツノゴケ属では、葉状体軸に沿って細い筋状の間隙が形成され、ここに藍藻が共生する。またツノゴケ科の葉状体中の細胞間隙は大きく発達している。
葉状体の中では、上記の細胞間隙を除いて組織・細胞の分化はほとんど見られない。ただしアナナシツノゴケ属などでは、表皮の細胞が明らかに小型である。胞子体と異なり、配偶体では、表皮の細胞も葉緑体をもつ。また粘液を多く含む細胞が葉状体中に散在している。造卵器の周囲などに細胞壁が階紋状に肥厚した細胞が見られることもある。
配偶体の先端 (縁辺部) には単一の頂端分裂細胞が存在し、先端成長をする。頂端細胞はふつう4個 (キノボリツノゴケ属では3個) の分裂面をもち、また周囲の細胞から分泌された粘液質に覆われて保護されている。
胞子体
ツノゴケ類の胞子体 (ゲノムを2セットもち胞子を形成する体) はその名のように細長い角状であり、ふつう配偶体から多数の胞子体が生じている (下図)。他のコケ植物とは異なり、ツノゴケ類の胞子体は柄を欠き、足と胞子嚢 (蒴) のみからなる。胞子嚢は活発な光合成を行い、長期間成長する。




足は球形であり、配偶体の背面に埋没している (右図)。足に接する配偶体の細胞では細胞壁が発達して表面積が大きくなり、胎座組織が形成される (他のコケ植物では足側の細胞でも細胞壁が発達する)。足の上には恒常性の分裂組織が存在し、その上部の胞子嚢に続いている (右図)。ツノゴケ類の胞子体は恒常性の分裂組織をもつという点で維管束植物の胞子体に類似しているが、この分裂組織が頂端分裂組織ではなく介在分裂組織 (分化した組織の間に挟まれた分裂組織) であるという点で異なる。ツノゴケ類の胞子体は恒常性の介在分裂組織があるため、長期間成長を続け、直立した胞子嚢先端から次第に成熟していく。ただしツノゴケモドキ属の胞子体では分裂組織が比較的早く分裂能を失い、横向きの短い胞子嚢が発生終期まで包膜内に留まっている (上図)。

胞子嚢 (蒴) は細長い角状であり、基部は配偶体由来の構造である包膜 (involucre) で包まれている (上図、右図)。配偶体とは異なり、胞子嚢の表皮細胞はふつう葉緑体を欠く。また表皮にはふつう2個の孔辺細胞で囲まれた気孔が存在し、孔辺細胞は葉緑体を含む。ツノゴケ類の気孔は基本的に開閉能を欠いており、胞子嚢を乾燥させるのが主な機能であると考えられている。またキノボリツノゴケ亜科やツノゴケモドキ属の胞子体は気孔を欠く。表皮の内側には葉緑体を含む同化組織 (assimilatory tissue) が存在し、その内側に胞子を形成する細胞、中央に軸柱 (columella) がある (左図)。胞子嚢は先端から順次成熟し、先端から2縦裂して胞子と偽弾糸 (pseudoelater) を放出する (上図)。苔類の弾糸と異なり、ツノゴケ類の偽弾糸はふつう明瞭な細胞壁のらせん状肥厚を欠き、またしばしば複数細胞からなる。
生活環

ツノゴケ類の生活環において、主となるのは単相の配偶体であり、接合子 (受精卵) から成長する複相の胞子体は配偶体上についている (右図)。ただし他のコケ植物に較べて、ツノゴケ類の胞子体は活発な光合成を行い、恒常性の分裂組織をもち (上記) 長期間成長するなど独立性が高い。胞子体は配偶体よりも高い光合成活性を示すことがあり、実験的には (胞子体だけに光を当てる) 胞子体から配偶体への光合成産物の輸送が起こる。


配偶体はふつう雌雄同株であるが、雌雄異株の種もいる。配偶子嚢 (造精器、造卵器) は配偶体の背面に埋まった状態で形成される (左図、右図)。
造卵器は腹部と頸部からなるフラスコ形であるが、配偶体背面にほとんど埋没している。この特徴は他のコケ植物とは異なり、維管束植物に類似している。造卵器の頸部は6列の細胞からなる。
配偶体背面の陥入部 (造精器腔 antheridial chamber) の底に、1〜80個の造精器が形成される (全て単一の細胞に由来する)。造精器の発生様式は、一部の苔類に類似した点がある。初期分岐群 (スジツノゴケ科、ツノゴケ科) では、造精器のジャケット細胞は4列にならぶが、その他では不規則に配列する。精子は細長く、巻いているが、他のコケ植物の精子と異なり、細胞構造は左右対称でねじれる方向が逆である。放出された精子は造卵器に達し、受精する。
接合子 (受精卵) は造卵器中に留まり、胞子体へと成長する。接合子の最初の分裂は縦方向に起こる (苔類や蘚類では横方向に分裂)。最初に足が形成され、その上部の分裂組織から胞子嚢が (蒴) が次第に形成されていく。発生初期の胞子嚢は配偶体に由来する包膜に包まれている。
ふつう胞子嚢は先端部から順次成熟し、胞子と偽弾糸を形成する (左図、右図)。胞子は減数分裂によって形成される単相の構造であるが、偽弾糸は複相の構造である。偽弾糸は乾湿運動 (水分量の変化によって変形する) を行い、胞子の効果的な散布に寄与すると考えられている。胞子嚢は先端部から2縦裂し、胞子と偽弾糸を放出する。このとき軸柱は残存する (右図)。
胞子の中には厚い黒褐色の細胞壁をもち20年以上休眠可能なものもあるし、薄い透明な細胞壁をもち短命なものもある。胞子壁の装飾や溝には多様性があり、重要な分類形質とされる。ふつう胞子は発芽後に細胞分裂するが、キノボリツノゴケ属では胞子内で細胞分裂して多細胞の状態で散布される。胞子は発芽して盤状または短い糸状の原糸体を形成する。原糸体はあまり発達せず、単一の葉状体を形成する。
他のコケ植物と同様、ツノゴケ類においても無性生殖が広く見られる。アナナシツノゴケ属の一種では、雄個体群と雌個体群が分断されており、無性生殖によってのみ個体群が維持されている。ツノゴケ類は、植物体の分断化や無性芽によって無性生殖を行う。
細胞
ツノゴケ類の細胞中には、ふつう1個の大きな葉緑体が存在し、葉緑体中にはルビスコ (光合成において二酸化炭素を固定する酵素) を主とするタンパク質の塊であるピレノイドが存在する。この特徴は陸上植物の中で特異であり、多くの緑藻に類似している。他の陸上植物では、ピレノイドを欠く盤状葉緑体が1細胞中に多数存在する。ピレノイドは、光合成における二酸化炭素濃縮機構に関与していると考えられている。ピレノイド基質にはふつう多数のチラコイドが侵入しており、またデンプン粒で囲まれている。ツノゴケ類の中で、ピレノイドの構造には多様性があり、分類形質となる。ただしスジツノゴケ属やアナナシツノゴケ属などはピレノイドを欠き、またアナナシツノゴケ属や Nothoceros は1細胞中に複数 (〜十数個) の葉緑体をもつ。ツノゴケ類の葉緑体では、チラコイドが管状の構造 (channel thylakoid) で垂直につながっている点でも他の陸上植物とは異なる。またツノゴケ類の細胞は、苔類に一般的な油体を欠く。
ネオクロム
ツノゴケ類は、陸上植物における重要な光受容体として知られるフィトクロムの一部とフォトトロピンのほぼ全長からなるキメラタンパク質であるネオクロム (neochrome) をもつ。この特異な光受容体は、緑色植物において接合藻と薄嚢シダ類、およびツノゴケ類に存在する。ネオクロムの遺伝子は、ツノゴケ類から薄嚢シダ類の多くを含む系統群の共通祖先に水平伝播したと考えられている。その年代は、およそ1億7900万年前と推定されている。
生態

他の多くのコケ植物と同様、ツノゴケ類は基本的に常緑多年生である。ツノゴケ類は、世界中の熱帯から温帯にかけて広く生育する。多くは湿った裸地などに生育するが、渓流沿いの岩上に生育するものや、生木の樹幹に着生しているものもいる (右図)。
上記のように、ツノゴケ類の葉状体内には、窒素固定能をもつネンジュモ属の藍藻が共生している。この共生藍藻はツノゴケ類と相利共生関係を結んでおり、固定した窒素化合物をツノゴケ類に供給し、ツノゴケ類から生育環境を提供してもらっている。
ツノゴケ類の配偶体には、ふつうグロムス類またはアツギケカビ類に属する菌根菌が共生している。また両者が同時に共生している例も少なくない。一方でキノボリツノゴケ属などは菌根菌を欠く。
系統
ツノゴケ類は、緑藻的な特徴 (ピレノイドを含む1個の葉緑体など)、苔類的な特徴 (造精器の発生様式など)、蘚類的な特徴 (軸柱をもつ胞子嚢など)、維管束植物的な特徴 (埋没した造卵器など) を併せもつことから、その系統的位置についてはさまざまな意見があった。また分子系統解析からも、さまざまな仮説が提唱されていた (下図)。
|
|
|
|
2000年代半ば以降、現生陸上植物の中で、苔類が最初に分岐し、次に蘚類が分岐、そしてツノゴケ類と維管束植物が姉妹群になる、という関係 (上図 3) が比較的広く受け入れられるようになった。この関係においては、ツノゴケ類と維管束植物に共通する形質 (埋没した造卵器、胞子体に恒常的な分裂組織、細胞壁にキシランが含まれる) は、両者の共有派生形質と考えることができる。
しかし2010年代半ば以降、より詳細な分子系統解析に基づき、コケ植物の3群 (苔類、蘚類、ツノゴケ類) が単系統群を構成しているとする仮説が支持されるようになった (上図 4)。コケ植物3群の中ではツノゴケ類が最初に分岐し、苔類と蘚類が姉妹群となる。この場合、ツノゴケ類と維管束植物に共通する形質は、現生陸上植物全体の共通祖先の形質であった可能性がある。
分類
世界中からおよそ300〜400種が報告されているが、実際の種数はそれよりも少なく、およそ150種ほどとされることが多い。日本からは6属17種が報告されている。
ツノゴケ類は、葉状性の配偶体をもつという外見的な類似性から、かつては苔類に分類されていた。しかし上記の様に特異な特徴を多くもつため、独立の分類群として扱われるようになった。1990年代ごろまではコケ植物門の1綱、ツノゴケ綱 (学名:Anthocerotopsida, Anthocerotae) として分類されていた。しかし21世紀にになるとコケ植物の単系統性が疑問視されることが多くなったためコケ植物門が使われなくなり、ツノゴケ類は独立の門、ツノゴケ植物門 (学名:Anthocerotophyta) に分類されることが多くなった。
20世紀中には、ツノゴケ類はふつう5〜6属ほどに分類されていた。また全ての種をツノゴケ目ツノゴケ科に分類する体系や、1目2科 (ツノゴケ科とツノゴケモドキ科) に分類する体系、2目3科 (ツノゴケ目ツノゴケ科とキノボリツノゴケ科、ツノゴケモドキ目・科) に分類する体系、4目5科 (ツノゴケ目ツノゴケ科とツノゴケモドキ科、ミヤベツノゴケ目・科、スジツノゴケ目・科、キノボリツノゴケ目・科) に分類する体系など、さまざまな分類体系が提唱されていた。
その後21世紀になると、分子系統学的研究によってツノゴケ類内の系統関係が次第に明らかとなった (下図)。その結果、既存の属が分割されて多数の属が提唱され、2019年現在では14属ほどに分けられることが多い。また、その系統関係に基づいて、ツノゴケ類を2綱5目5科に分類する体系が提唱されている (下表)。
| |||||||||||||||||||||||||||
| ツノゴケ類の系統仮説. |
ツノゴケ類の分類体系の1例 (2019年現在)
|
ギャラリー

ツノゴケ類の一種

ツノゴケ属の一種

ニワツノゴケ属の一種
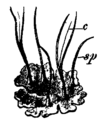
模式図 (c = 軸柱、sp = 胞子嚢)




脚注
注釈
関連項目
外部リンク
- コケ植物・ツノゴケ. 植物形態学. 福原のページ(植物形態学・生物画像集など).
- Glime, J. M. (2017) Anthocerotophyta. Chapt. 2-8. In: Glime, J. M. Bryophyte Ecology. Volume 1. Physiological Ecology. Ebook 2-8-1 sponsored by Michigan Technological University and the International Association of Bryologists. (英語)
- Phylum Anthocerotophyta. Introduction to Bryophytes. Biology 321, University of British Columbia. (英語)
- What is a hornwort? Australian Bryophytes. The Australian National Botanic Gardens. (英語)