

細胞表面受容体 (cell surface receptors, または膜受容体: membrane receptors、膜貫通受容体: transmembrane receptors) は、細胞の原形質膜に埋め込まれた受容体である。それらは細胞外分子を受け取る (結合する) ことで、細胞シグナル伝達に作用する。それらは、細胞と細胞外空間との間のコミュニケーションを可能にする特殊な内在性膜タンパク質である。細胞外分子は、ホルモン、神経伝達物質、サイトカイン、成長因子、細胞接着分子、または栄養素などであり、受容体と反応して細胞の代謝や活性の変化を引き起こす。シグナル伝達の過程では、リガンド結合は、細胞膜を介した生化学的カスケードに影響を与える。
構造と機構
膜受容体の多くは膜貫通型タンパク質である。糖タンパク質やリポタンパク質など様々な種類がある。何百種類もの受容体が知られており、さらに多くの種類がまだ研究されていない。膜貫通型受容体は、通常、その三次構造 (三次元構造) に基づいて分類される。三次元構造が不明な場合は、膜トポロジーに基づいて分類できる。最も単純な受容体では、ポリペプチド鎖は脂質二重層を一度だけ横断するが、Gタンパク質共役受容体のような受容体は7回も横断する。各細胞膜は、さまざまな表面分布を持ついくつかの膜受容体を持つことができる。単一の受容体は、膜の種類や細胞機能に応じて、異なる膜位置に異なって分布することもある。受容体は、膜表面に均等に分布しているのではなく、膜表面に集まることが多くある。
機構
膜貫通型受容体の作用メカニズムを説明するために、2つのモデルが提案されている。
- 二量体化 (dimerization): 二量体化モデルは、リガンド結合の前に、受容体が単量体の形で存在することを示唆している。アゴニスト結合が起こると、モノマーが結合して活性二量体を形成する。
- 回転 (rotation): 受容体の細胞外部分に結合するリガンドは、受容体の膜貫通ヘリックスの一部の回転 (コンホメーション変化) を誘発する。回転により、受容体のどの部分が膜の細胞内側に露出しているかが変わり、受容体が細胞内の他のタンパク質と相互作用する方法が変わる。
ドメイン
原形質膜の膜貫通型受容体は、通常3つの部分に分けることができる。
細胞外ドメイン
細胞外ドメインは、細胞または細胞小器官のすぐ外側にある。ポリペプチド鎖が二重層を数回横断する場合、外部ドメインは、膜を通して絡み合ったループからなる。定義により、受容体の主な機能は、ある種のリガンドを認識して応答することである。例えば、神経伝達物質、ホルモン、または原子イオンは、それぞれ、受容体に結合したリガンドとして細胞外ドメインに結合することができる。クロトー (英語版) は、受容体に作用してリガンド(FGF23 (英語版) )を認識する酵素である。
膜貫通ドメイン
膜貫通型受容体の最も豊富なクラスは、GPCRとシングルパス膜貫通型タンパク質 (英語版) である。ニコチン性アセチルコリン受容体などの一部の受容体では、膜貫通ドメインが膜を貫通して、またはイオンチャネルの周囲にタンパク質の細孔を形成する。適切なリガンドの結合によって細胞外ドメインが活性化されると、細孔はイオンにアクセスできるようになり、イオンは拡散する。他の受容体では、膜貫通ドメインは結合するとコンホメーション変化を起こし、細胞内の状態に影響を与える。7回膜貫通型受容体(7TM)スーパーファミリーのメンバーなどの一部の受容体では、膜貫通ドメインにリガンド結合ポケットが含まれている。
細胞内ドメイン
受容体の細胞内 (または細胞質) ドメインは、細胞や細胞小器官の内部と相互作用し、シグナルを中継する。この相互作用には2つの基本的な経路がある。
- 細胞内ドメインは、エフェクター・タンパク質とのタンパク質-タンパク質相互作用を介してシグナルを伝達し、エフェクター・タンパク質はシグナルを宛先に伝達する。
- 酵素結合型受容体では、細胞内ドメインに酵素活性を持っている。多くの場合、これはチロシンキナーゼ活性である。酵素活性は、細胞内ドメインに関連する酵素に起因することもある。
シグナル伝達

膜受容体を介したシグナル伝達プロセスには、リガンドが膜受容体に結合する外部反応と、細胞内反応が引き起こされる内部反応が含まれる。
膜受容体を介したシグナル伝達には、次の4つの部分が必要である。
- 細胞外シグナル伝達分子:細胞外シグナル伝達分子は、1つの細胞で産生され、少なくとも隣接する細胞に移動することができる。
- 受容体タンパク質:細胞は、シグナル伝達分子に結合して細胞内に伝達する細胞表面受容体タンパク質が必要である。
- 細胞内シグナル伝達タンパク質:これらは細胞小器官にシグナルを渡す。シグナル分子が受容体タンパク質に結合すると、細胞内シグナル伝達タンパク質が活性化され、シグナル伝達カスケードを開始する。
- 標的タンパク質:シグナル伝達経路が活性化され、細胞の挙動が変化すると、標的タンパク質のコンホメーションや他の特性が変化する。

膜受容体は主に構造と機能によって3つのクラスに分けられる。イオンチャネル結合型受容体 (英語版) 、酵素結合型受容体 (英語版) 、Gタンパク質共役型受容体である。
- イオンチャネル結合型受容体は、アニオンとカチオンのイオンチャネルを持ち、マルチパス膜貫通型タンパク質の大きなファミリーを構成している。それらは通常、ニューロンなどの電気的に活性な細胞で見られる急速なシグナル伝達イベントに参加している。それらはリガンド依存性イオンチャネルとも呼ばれている。イオンチャネルの開閉は神経伝達物質によって制御される。
- 酵素結合型受容体は、酵素そのものか、関連する酵素を直接活性化する。これらの受容体は通常、1回膜貫通型受容体であり、受容体の酵素成分は細胞内に保持されている。酵素結合型受容体の大部分は、プロテインキナーゼであるか、またはプロテインキナーゼと結合している。
- Gタンパク質共役型受容体は、7回膜貫通型ヘリックスを持つ膜内在性タンパク質である。これらの受容体は、アゴニストが結合するとGタンパク質を活性化し、Gタンパク質は細胞内シグナル伝達経路における受容体効果を仲介する。
イオンチャネル連結型受容体
ニューロンの信号伝達イベント中に、神経伝達物質は受容体に結合し、タンパク質のコンホメーションを変化させる。これによりイオンチャネルが開き、細胞外イオンを細胞内に入れることができる。原形質膜のイオン透過性が変化し、これにより細胞外の化学信号が細胞内の電気信号に変換され、細胞の興奮性 (英語版) を変化させる。
アセチルコリン受容体は、カチオンチャネルに連結された受容体である。4つのサブユニット (α、β、γ、δサブユニット) から構成されている。αサブユニットは2つあり、それぞれアセチルコリン結合部位が1つある。この受容体は、3つのコンホメーションで存在することができる。閉状態-非占有状態は本来のタンパク質の構造である。アセチルコリンの2つの分子が両方ともαサブユニット上の結合部位に結合すると、受容体のコンホメーションが変化し、ゲートが開いて、多くのイオンや低分子の侵入が可能になる。ただし、この開状態-占有状態はわずかな期間しか持続せず、その後ゲートが閉じられ、閉状態-占有状態になる。アセチルコリンの2つの分子は、すぐに受容体から解離し、本来の閉状態-非占有状態に戻る。
酵素結合型受容体
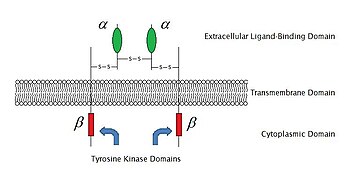
2009年現在、6種類の酵素結合型受容体が知られている。
受容体チロシンキナーゼは、最も多くの個体数を有し、最も広範な応用が可能である。これらの分子の大部分は、上皮成長因子(EGF)、血小板由来成長因子 (PDGF)、線維芽細胞成長因子 (FGF)、肝細胞増殖因子(HGF)、神経成長因子(NGF)、およびインスリンなどのホルモン等成長因子の受容体である。これらの受容体のほとんどは、さらなるシグナル伝達を活性化するために、それらのリガンドと結合した後に二量体化する。例えば、上皮成長因子(EGF)受容体がそのリガンドEGFと結合した後、2つの受容体は二量体化し、その後、各受容体分子の酵素部分のチロシン残基のリン酸化を受ける。これによりチロシンキナーゼが活性化し、さらに細胞内反応を触媒する。
Gタンパク質共役受容体
Gタンパク質共役型受容体は、膜貫通型受容体の大規模なタンパク質ファミリーを構成している。これらは真核生物にのみ存在する。これらの受容体に結合し活性化するリガンドには、感光性化合物、臭気、フェロモン、ホルモン、神経伝達物質などがある。これらのリガンドの大きさは、低分子からペプチドや大型タンパク質まで様々である。Gタンパク質共役型受容体は多くの疾患に関与しているため、現代の多くの医薬品の標的となっている。
Gタンパク質共役型受容体が関与する2つの主要なシグナル伝達経路がある:cAMPシグナル伝達経路とホスファチジルイノシトールシグナル伝達経路である。どちらもGタンパク質の活性化を介して媒介される。Gタンパク質は、α、β、およびγとして指定された3つのサブユニットを持つ三量体タンパク質である。受容体の活性化に応答して、αサブユニットは、結合したグアニシン二リン酸(GDP)を放出し、それはグアニシン三リン酸(GTP)によって置換され、それによりαサブユニットを活性化し、βおよびγサブユニットから解離する。活性化されたαサブユニットは、さらに細胞内シグナル伝達タンパク質または標的機能タンパク質に直接影響を与えることができる。
膜受容体関連疾患
膜受容体が変性または欠損していると、シグナル伝達に支障をきたし、疾患を引き起こす可能性がある。一部の疾患は、膜受容体機能の障害によって引き起こされる。これは、受容体タンパク質をコードし、調節する遺伝子の変化による受容体の欠損や変性が起こるためである。膜受容体 TM4SF5は、肝細胞の遊走や肝腫に影響を与える。また、皮質NMDA受容体は膜の流動性に影響を与え、アルツハイマー病では変化する。細胞がエンベロープのないウイルスに感染すると、ウイルスはまず特定の膜受容体に結合し、その後、それ自体またはサブウイルス成分を細胞膜の細胞質側に通過させる。ポリオウイルスの場合、受容体との相互作用がコンホメーションの再配列を引き起こし、VP4と呼ばれるビリオンタンパク質を放出することがin vitroで知られている。VP4のN末端はミリスチル化されているため、疎水性である (ミリスチン酸=CH3(CH2)12COOH)。受容体の結合によって誘導されるコンホメーション変化により、VP4にミリスチン酸が付着し、RNAのチャネルが形成される結果となることが提案されている。
構造に基づいた医薬品設計

X線結晶構造解析やNMR分光法などの手法により、標的分子の3次元構造に関する情報が飛躍的に増加し、リガンドの構造情報が得られるようになったことで、構造に基づいた医薬品設計の開発が急速に進んでいる。これらの新薬の中には膜受容体を標的としたものもある。構造に基づいた医薬品設計の現在のアプローチは、2つのカテゴリーに分類できる。第1のカテゴリーは、特定の受容体のリガンドを決定することに関するものである。これは通常、データベースクエリ、生物物理学的シミュレーション、および化学ライブラリーの構築によって行われる。いずれの場合も、多数の潜在的なリガンド分子をスクリーニングして、受容体の結合ポケットに適合するものを見つけ出す。このアプローチは通常、リガンドベースのドラッグデザインと呼ばれている。データベースを検索することの主な利点は、新しい有効な化合物を得るための時間と労力を節約できることである。構造に基づく医薬品設計のもう一つのアプローチは、リガンドを組み合わせてマッピングすることであり、これは受容体に基づく医薬品設計と呼ばれている。この場合、リガンド分子は、小さな断片を段階的に組み立てることにより、結合ポケットの制約内で設計される。これらの断片は、原子または分子のいずれでもよい。このような方法の主な利点は、新しい構造を発見できることである。
その他の例
- アドレナリン受容体
- 嗅覚受容体
- 受容体チロシンキナーゼ
- 上皮成長因子受容体
- インスリン受容体
- 線維芽細胞成長因子受容体 (英語版)
- 高親和性ニューロトロフィン受容体 (英語版)
- エフリン受容体
- インテグリン
- 低親和性神経成長因子受容体 (英語版)
- NMDA受容体
- いくつかの免疫受容体 (英語版)
関連項目
- セカンドメッセンジャー
- ニューロモデュレーション (神経調節) (英語版)
参考文献
外部リンク
- IUPHAR GPCR データベース
- Cell Surface Receptors - MeSH・アメリカ国立医学図書館・生命科学用語シソーラス(英語)