
原文と比べた結果、この記事には多数の(または内容の大部分に影響ある)誤訳があることが判明しています。情報の利用には注意してください。
|

進化発生生物学 (しんかはっせいせいぶつがく、英: Evolutionary developmental biology、通称: evo-devo〈エボデボ〉とも)は、異なる生物の発生過程を比較してそれらの系統関係を推測し、発生過程がどのように進化したかを示す生物学的研究の分野である。
この分野は、19世紀の初めに誕生した。当時、動物学者は、胚の発生が分子レベルでどのように制御されているかを知らなかった。チャールズ・ダーウィンは、似た胚を持つことは共通の祖先を暗示していると指摘したが、1970年代までほとんど進展がなかった。その後、組換えDNA技術が分子遺伝学と同時に発生学に大きな発見をもたらした。初期の重要な発見は、広範囲の真核生物における発生を制御するホメオティック遺伝子の発見であった。
この発見はいくつかの重要な概念によって特徴づけられ、生物学者を驚かせるものであった。
ひとつは深い相同性である。長い間別々に進化してきたと考えられている昆虫、脊椎動物および軟体動物の「目」の発生は、pax-6のような同じ遺伝子によって制御されていることがわかった。pax-6は転写因子だが、こういった転写因子、傍分泌因子、シグナル伝達カスケードの遺伝子は「ツールキット遺伝子」といわれ、古代から受け継がれており、門を越えて高度に保存されている。それらは胚を形作る時間と空間のパターンを生成し、最終的には生物の体を形成する。
もうひとつは、酵素のような構造遺伝子は、種の間であまり変わらない。異なるのは、遺伝子発現がツールキット遺伝子によって調節される方法である。これらの構造遺伝子は、胚の異なる部分で、異なる発生段階で何度も再利用され、制御の複雑なカスケードを形成し、他の調節遺伝子および構造遺伝子を正確なパターンでオンオフする。
新しい形態学的特徴および最終的には新しい生物の「種」ができるのは、ツールキットによってであり、ツールキット遺伝子が新しいパターンで発現される場合、またはツールキット遺伝子がさらなる機能を獲得する場合のいずれかにおいてである。もう一つの可能性はエピジェネティックな変化が後に遺伝子レベルで統合されるという新ラマルク説である。これは、多細胞生命の歴史の初期において重要であったかもしれない。
歴史

反復説
1824-1826年にエティエンヌ・セールは進化発生の反復説を提唱したが、アリストテレスによって着想を得たヨハン・フリードリヒ・メッケルの1808年の考えが反映されている。彼らは、「より高等な」動物の胚の各段階は、「生物の大きな連鎖」の下の段階の動物の一連の段階を経る、つまり反復されると主張した。例えば、人間の胚は、最初は魚の胚ようにみえ、次に爬虫類、鳥類、哺乳動物とよく似た段階を経て、人間の胚になる、という観察にもとづく。発生学者のカール・エルンスト・フォン・ベーアは、1828年に、単一のボディープランに基づいて、「生物の大きな連鎖」のような線形の順序はないと主張し、これに反対した。フォン・ベーアは代わりに、4つの異なる動物のボディプランを提唱した。ヒトデのような放射状の動物、貝類のような軟体動物、ロブスターのような甲殻の動物、魚のような脊椎動物。その後、動物学者たちは、1866年にエルンスト・ヘッケルが反復説を唱えたにもかかわらず、概ねこの理論を放棄した。
進化形態学

19世紀初頭から20世紀まで、発生学は謎と対峙していた。動物は、卵からしばしば同様の段階を経て、大きく異なる体制の成体になるように見えるが、動物学者は胚発生が分子レベルでどのように制御されているかについてほとんど何もわかっていなかった。チャールズ・ダーウィンは、共通の胚構造が共通の祖先を暗示していると主張した。その一例として、ダーウィンは、1859年の著作『種の起源』に、エビに似たフジツボの幼虫を引用し、その成虫が他の節足動物のようには見えないと、指摘した。リンネとキュヴィエはそれらを軟体動物に分類した。19世紀の動物学は、発生学を進化科学に変え、胚の相同性を系統発生に結びつけた。フリッツ・ミューラーを含む動物学者は、分類群間の系統関係を発見するための発生学を提案した。しかし、次の100年の間、分子的証拠がなく、進展は止まった。
evo-devoの誕生
1977年、遺伝学における組換えDNA技術の到来により、進化と発生生物学の考え方の革命が始まり、スティーヴン・ジェイ・グールドのオントロジーに関する論文『個体発生と系統発生』とフランソワ・ジャコブにより進展が始まった。グールドはヘッケルの進化論的発生学の解釈を退け、ジャコブは別の理論を立てた。これは、最終的に、発生学および分子遺伝学、系統発生、進化生物学を含めて、evo-devoの基礎をつくった。1978年、エドワード・B・ルイスはショウジョウバエの胚の発育を調節するホメオティック遺伝子を発見した。昆虫が含まれる無脊椎動物の主要な門の一つである節足動物に属するが、Bill McGinnisは、カエル、鳥、哺乳類などの脊椎動物や、他の門の動物で、ホメオティック遺伝子に共通の配列、ホメオボックスを発見した。ホメオボックスは後に酵母や植物にも見出された。すべての真核生物にわたって発生を制御する遺伝子に明らかに強い類似性があった。1980年、クリスティアーネ・ニュスライン=フォルハルトとエリック・ヴィーシャウスは、ショウジョウバエの胚にセグメンテーションパターンを作り出すのに役立つギャップ遺伝子を発見した。彼らとルイスは1995年にノーベル賞を受賞した。その後、より特異的な類似性が発見された。例えば、Distal-less遺伝子は、ショウジョウバエの付属肢や四肢の発生に関与することが1989年に発見されただけでなく、魚のヒレ、ニワトリの羽、海洋性の環形動物の足(疣足)、ホヤの出水口、ウニの管足などの発生に関与することがわかった。その遺伝子を動物が獲得したのは古代だと考えなければならず、はなれた動物の共通の祖先をたどると、エディアカラ時代の前、約6億3,500万年前に獲得されたと考えないといけないことは明らかであった。Evo-devoは、すべての動物の体が発生中にどのようにつくられるのかを解明し始めた。
体の構造の制御
深い相同性
動物の卵は、クラゲからロブスター、蝶からゾウまで、非常に異なる体を生じさせる。これらの生物の多くは、コラーゲンや酵素のような身体を構成するタンパク質においては、同じ遺伝子を共有しているが、生物学者は各動物群に独自の発生のルールがあると予想していた。エボ・デボの驚きは、さまざまな動物の種によるの体の違いに寄与するのは、遺伝子が異なるからではなく、調節遺伝子すらも、すべての動物が古代から共有してきたものであったということである。キリンは長い頸部の遺伝子を持っていない。象は大きな体の遺伝子を持っていない。彼らの体の発生は、切り替えシステムによって異なっていた。つまり、いつはじめるか、胚のこの部分またはその部分に生じさせるか、発現の時間を多くするか少なくするかなどにより、パターン化され異なる特徴が発生で生じる。
胚発生が制御される方法の謎は、ショウジョウバエをモデル生物として解決され始めた。その胚発生の制御は、蛍光色素を、胚で発現されるタンパク質に結合させ、視覚化された。目的のタンパク質が、生きている胚の、どこにいつ出現するかが調べられた。

このような技術を用いて、1994年にヴァルター・ゲーリングは、ショウジョウバエの目を形成するために不可欠なpax-6遺伝子が、マウスおよびヒトの眼形成遺伝子と正確に一致することを見出した。同じ遺伝子は、イカなど、軟体動物頭足類のような他の多くの動物群ですぐに見つかった。Ernst Mayrを含む生物学者は、異なる種類の眼の解剖学的構造の違いがとても大きいので、少なくとも40回は動物界で目が別々に進化してきたと信じていた。例えば、ショウジョウバエの複眼は、何百もの小さなレンズ構造(個眼鏡)で作られている。人間の目には視神経が目に入る盲点があり、神経線維は網膜の表面に行き渡るので、網膜の視神経に到達する前に光が神経線維の層を通過しなければならないので、構造的に両者は "逆さま"である。対照的に、軟体動物頭足類の眼には網膜があり、次に神経線維の層があり、次に眼の壁があり、「まっすぐ」である。つまり、2つの眼の構造はまったく逆になっている。しかし、pax-6が共通していることは、同じ遺伝子がすべての動物の目の発生を制御し、それらがすべて共通の祖先から進化したことを示唆している。古代の遺伝子が、何百万年もの進化によって保存され、類似の機能をもって、異種間の構造をつくり出しているということは、かつて機能が似ているだけと考えられていた構造間にある、深い相同性を示している。これは進化生物学における相同性の意味の根本的な改訂を引き起こしている。
遺伝子ツールキット

生物のゲノム中の少数の遺伝子が生物の発生を制御する。これらの遺伝子は、発生に関わるツールキット遺伝子と呼ばれている。それらは門の間で高度に保存されており、進化の初期で獲得されたことを意味している。ツールキット遺伝子は、発生において、ボディプランと、身体部分の数、同一性、パターンの違いに影響を及ぼす。ほとんどのツールキット遺伝子は、シグナル伝達経路の一部であり、転写因子、細胞接着タンパク質、細胞表面受容体タンパク質およびそれらに結合するシグナル伝達リガンド、ならびに胚を通って拡散する分泌されたモルフォゲンをコードする。これらのすべては、胚の未分化細胞の運命を決定するのに役立つ。それと同時に、ツールキット遺伝子は、胚を形作る、時間と空間のパターンを生成し、最終的に生物のボディプランを形成する。最も重要なツールキット遺伝子の中にHox遺伝子がある。これらの転写因子は、他のツールキット遺伝子にも見出されるホメオボックスタンパク質結合DNAモチーフを含み、その前後の軸に沿って身体の基本パターンを生成する。Hox遺伝子は、ヘビの多くの脊椎のような反復部分が発育中の胚または幼虫でどのくらい成長するかを決定する。既に言及したPax-6は、古典的なツールキット遺伝子である。ホメオボックス遺伝子は、植物においても見出され、それらは全ての真核生物に共通であることを示唆している。
胚の制御ネットワーク

ツールキットのタンパク質産物は、重複および改変ではなく、多面的なモザイクによって再利用され、多くの独立した発生過程で不変的に適応され、多くの異なる身体構造にパターンを与える。ツールキット遺伝子は、大きく複雑な調節配列をもつ。例えば、ショウジョウバエのロドプシン遺伝子は、わずか数百塩基対の長さの調節配列を有するが、アイレスシス遺伝子の調節領域は、7000塩基対以上に6つのシス調節エレメントを含む。関与する制御ネットワークはしばしば非常に大きい。各調節タンパク質は、数百の調節配列を制御する。例えば、ショウジョウバエの67個の転写因子は、それぞれ平均124個の標的遺伝子を制御する。この複雑さはすべて、胚の発生に関与する遺伝子を、正確なタイミングで、正確な場所で、正確にスイッチをオン/オフすることを可能にする。スイッチをオンオフされる遺伝子のいくつかは、構造遺伝子であり、酵素をつくったり、胚の組織および器官を直接形成する。しかし、他の多くの遺伝子はそれ自体が調節遺伝子であるため、スイッチを入れることはしばしば正確にタイミングを取った切替えのカスケードであり、発生中の胚で別の発生プロセスを開始することになる。

このようなカスケード調節ネットワークは、ショウジョウバエの胚の発生において詳細に研究されている。若い胚は、ラグビーボールのような形の楕円形である。少数の遺伝子が、胚の長軸に沿って濃度勾配をつくり出す。初期胚では、ビコイドbicoidおよびハッチバックhunchback遺伝子が、前端付近で高濃度であり、将来の頭および胸郭にパターンを与える。コーダル caudal やナノスnanos遺伝子は後端付近で高濃度であり、後ろの腹部にパターンを与える。これらの遺伝子の作用は相互作用する。例えば、Bicoidタンパク質は尾部のメッセンジャーRNAの翻訳をブロックするので、尾部タンパク質濃度は前端部で低くなる。コーダルは後ろ側でハエの後肢部分を生成する遺伝子をスイッチを入れるが、後端部で最も集中している。
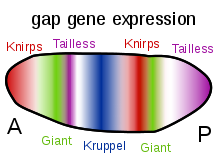
ビコイドBicoid、ハッチバックHunchbackおよびコーダルCaudalタンパク質は、giant遺伝子、knirps遺伝子、Kruppel遺伝子、tailless遺伝子などのギャップ遺伝子の転写を、縞状のパターンの発現で制御し、セグメントとなる構造の最初の段階をつくり出す。これらのタンパク質は、次に、ペアルールpair-rule遺伝子を制御し、次の段階で、胚の長軸に7つのバンドをつくり出す。最後に、刻み目のようなセグメントポラリティ segment polarity遺伝子は、7つのバンドのそれぞれを2つに分割し、14のセグメントをつくり出す。
このプロセスは、異なる種の動物において、ツールキット遺伝子は深い相同性をもち、つまり塩基配列が正確に保存されており、同じ機能をもっていることを説明する(例えば、マウスのタンパク質がショウジョウバエの発育を制御でき、その相同性がわかる)。転写因子とシス調節エレメント、またはシグナル伝達タンパク質と受容体の相互作用は、複数の用途で使われるため、ほぼすべての変異が有害であるため、変わらず保存されるのである。
新規性はどこから?
より驚くべき、そしておそらく(新ダーウィンの観点から)直観的ではない進化発生生物学における最近の研究の結果は、多くの門を越えた生物体の形態の多様性が、発生に関連する高度に保存された、ツールキット遺伝子やその他の遺伝子によっており、それらの遺伝子の塩基配列の多様性によっていないことである。確かに、GerhartとKirschnerが指摘しているように、明らかな矛盾がある。「変化が最も期待される場所では、変化がなく、保存がされていることが確認されるだけである」。異なる動物の間で観察された形態学的な新規性が(変異などによる)遺伝子配列の変化に由来しない場合、いったい、何に由来するのだろうか?
それは、新規性は、遺伝子調節が、突然変異によって変化することによって、生じ得る。
ツールキットのバリエーション
ツールキットのバリエーションは、動物の形態学的進化の大部分を生み出した可能性がある。ツールキットは2つの方法で進化を推進することができる。ツールキット遺伝子は全く違うパターンで発現されうる。例えば、ダーウィンフィンチの種によって異なるくちばしの大きさが、BMP遺伝子の発現量に関連することまたはdistal-less遺伝子の発現が減り、ヘビが脚を失ったことなどが、挙げられる。あるいは、ツールキット遺伝子は、新しい機能を獲得することができる。例えば、同じdistal-less遺伝子には、多くの機能があり、脊椎動物の下顎のような多様な構造を制御し、ショウジョウバエの足、触角を形成し、蝶の翅の目のような模様のパターンをつくる。
エピジェネティックな変化の統合
進化は、ときにラマルク説にあうように、始まるのかもしれない。エピジェネティックな変化により、遺伝子調節または表現型の変化が起こり、その後、遺伝子レベルでの変化によって統合される。エピジェネティックな変化には、可逆的メチル化によるDNAの改変を含み、発生の機構がもつ遺伝的な可塑性により、肉体的および他の環境効果によって、生物のプログラムされていない変化が含まれる。生物学者Stuart A. NewmanとGerd B. Mullerは、多細胞生物の歴史の初期の生物は、エピジェネティックな進化に対してより強く反応して、初期の大進化の基礎を提供したかもしれないと示唆している。
エコエボデボ eco-evo-devo
生態-進化発生生物学 (eco-evo-devo) は、進化論との関係を調べるために、発生生物学と生態学の研究を統合する。研究者は、発生的可塑性、後成的遺伝、遺伝的同化、ニッチ構築および共生などの概念および機構を研究する。
脚注
参考文献
- Secord, James A. (2000), Victorian sensation: the extraordinary publication, reception, and secret authorship of Vestiges of the natural history of creation, Chicago: University of Chicago Press, ISBN 978-0-226-74410-0