
| 霊長目 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

バーバリーマカク Macaca sylvanus
| ||||||||||||||||||
| 保全状況評価 | ||||||||||||||||||
| ワシントン条約附属書II | ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Primates Linnaeus, 1758 | ||||||||||||||||||
| 和名 | ||||||||||||||||||
| 霊長目 | ||||||||||||||||||
| 亜目 | ||||||||||||||||||
霊長目(れいちょうもく、Primates)は、哺乳綱に分類される目。別名サル目、霊長類。
生物学的には、ヒトも霊長目の一員であり、霊長類(=サル類)の一種にほかならないが、一般的には、霊長目からヒトを除いた総称を「サル」とする。
分布
(以下の記述はヒトを除いたサル目の種に関するものである)
熱帯系の動物であり、その分布は熱帯域に集中する。東アジアには温帯域まで分布する種があり、特にニホンザルは最も高緯度に分布するサルとして有名である。曲鼻亜目及びメガネザル類はアジア・アフリカの熱帯域、広鼻猿類は中南米の熱帯、類人猿を含む狭鼻猿類はアジア・アフリカの熱帯域から温帯域の一部にかけて分布している。ヨーロッパにはほとんど棲息せず、ジブラルタル海峡ごしにバーバリーマカク1種が棲息するのみである。また、北アメリカ大陸北部(アングロアメリカ相当地域)にも分布しない。
形態
有蹄類を除く哺乳類や鳥類は左右が湾曲した筒状になる鋭い爪(鉤爪, claw)があるが、本目の構成種の指趾には扁平な爪(平爪, nail)がみられる。平爪は指趾の先端から大きくは伸びないため、指先の器用さを確保することができる。曲鼻類は後肢の第2趾に鉤爪があり、毛づくろいなどに用いる。例外はアイアイで後肢の第1趾のみ平爪があり、他の指趾には鉤爪がある。直鼻類ではマーモセット類も後肢の第1趾のみ平爪があり、他の指趾には鉤爪がある。
体重100g以下のコビトガラゴ Galagoides demidovii から、200kgを超すゴリラまで、多様な種が属している。
サル目は、哺乳類としては比較的基本的な体制を維持している。爪や歯などには大きな特殊化は起こっていない。その中で、サル類を特徴づけるのは、以下のような点である。
- ほとんどの種では手と足に5本の指をもち、手と足の親指が他の4本と多少とも対向しているため、物をつかむことができる。このことから、霊長類はかつては(足が二足歩行に適した形に進化した人類に対して)「四手類(Quadrumana)」と呼ばれた。
- 両目が顔の正面に位置しており、遠近感をとらえる能力に優れている。
- 直鼻亜目 Simiiformes 下目 Catarrhini 小目(狭鼻猿類)は3色型色覚を有し、緑色の葉の間から、さまざまな色をした果実などを見つけるのに有利になっている。その他の霊長類は特にオスで2色型色覚にとどまっている種が大半である。色覚の詳細については後述する。
また、
生態
曲鼻猿類はキツネザル類に昼行性が多いのを除けば夜行性がほとんどだが、直鼻猿類はメガネザル類と広鼻猿に属するヨザル類を除いてほぼ全てが昼行性である。生活環境は樹上生活から地上生活まで幅広い。
食性も昆虫食、果実食、草食など、多岐にわたる。ただし、全体としてみれば、樹上性のものが多い。地上性のものはそこから派生したと考えられる。
分類
「霊長」という言葉において、霊は魂や幽霊という漢字そのものの意味より、優れたもの、不思議な力を持っているという意味が強い。つまり、これはヒトや、ヒトを含むサルの仲間を、動物の進化の最終形態とする認識から付けられた名前である。英語名の Primate も、大主教や最高位を意味する単語であり、やはり同様の観点から付けられた名前である。
以前は主に脳が小型で嗅覚が発達し鼻面の長いキツネザル類・ロリス類・メガネザル類を原猿亜目 Prosimii、それ以外の主に脳が大型で視覚が発達し鼻面の短い分類群を真猿亜目 Anthropoidea としてまとめていた。研究の進展により、メガネザルがいわゆる原猿類の他のグループよりも真猿類により近いことが判明した。このことから、現在ではキツネザル類・ロリス類をまとめて「曲鼻猿類(曲鼻猿亜目、曲鼻類、曲鼻亜目)」、メガネザル類を含むその他の霊長類を「直鼻猿類(直鼻猿亜目、直鼻類、直鼻亜目)」と呼び、正式な分類体系では、「原猿類」という名称は用いなくなっている。

以下の分類・和名は、日本モンキーセンター霊長類和名編纂ワーキンググループ (2018)に従う。ただしほとんどの和名は共通するが、下目の和名については川田ら(2018)に従った。
-
直鼻亜目 Haplorrhini
- メガネザル型下目 Tarsiiformes
-
真猿型下目 Simiiformes
-
狭鼻小目 Catarrhini
- オナガザル上科 Cercopithecoidea
- ヒト上科 Hominoidea
- 広鼻小目 Platyrrhini
-
狭鼻小目 Catarrhini
- 曲鼻亜目 Strepsirrhini
進化
|
真主齧上目 Euarchontoglires |
|
||||||||||||||||||||||||||||||
霊長類の最古の化石は、白亜紀末期の北アメリカ西部から発見されており、プレシアダピス類(偽霊長類)と呼ばれる。このように、霊長類の進化は約6,500万年前、白亜紀末期頃に始まったと考えられている。
新生代に入り暁新世になるとアダピス類とオモミス類が繁栄した。いずれもまだ原始的な種類で、アダピス類は後の曲鼻猿類に、オモミス類が直鼻猿類に進化したと考えられる。直鼻亜目と曲鼻亜目の分岐と同時期の6,300万年前に、直鼻亜目はL-グロノラクトンオキシダーゼ(ビタミンC合成酵素)の酵素活性を失っている。 アダピス類とオモミス類はヨーロッパと北アメリカに分布したが、何らかの環境要因によって北アメリカの霊長類は絶滅し、以降、ユーラシアとそれに近接していたアフリカという旧世界の大陸を舞台に霊長類の進化は進んだ。曲鼻亜目の一部は海によって他の大陸から隔絶されていたマダガスカル島にアフリカから進出し(恐らくは流木等に掴まっての漂着)、キツネザル類に進化していった。
その後、直鼻亜目が中新世にはアジア・アフリカに住む狭鼻猿類と南アメリカの広鼻猿類とに分かれる。上述のように北アメリカの猿類は絶滅したので、南米の広鼻猿類の祖先はアフリカから渡って来たとの説が有力である(当時、アフリカ大陸と南米大陸は既に分裂していたが、両大陸間の大西洋は現在と比較すれば狭く、距離は近かった。そのため小型の猿類ならば流木等を使って漂着できた可能性がある)。広鼻猿類の祖先やテンジクネズミ上科の祖先がアフリカでできた浮島に乗って大西洋を流されて南米大陸に到着したという説も紹介されている。真猿下目の狭鼻猿類(旧世界ザル)と広鼻猿類(新世界ザル)とが分岐したのは3,000-4,000万年前と言われている。
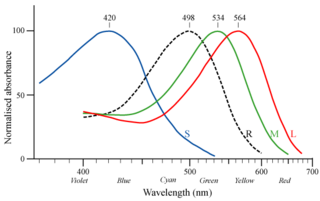
脊椎動物の色覚は、網膜の中にどのタイプの錐体細胞を持つかによって決まる。魚類、両生類、爬虫類、鳥類には4タイプの錐体細胞(4色型色覚)を持つものが多い。よってこれらの生物は長波長域から短波長域である近紫外線までを認識できるものと考えられている。一方ほとんどの哺乳類は錐体細胞を2タイプ(2色型色覚)しか持たない。爬虫類の祖先から枝分かれした哺乳類の祖先は当初は4タイプ全ての錐体細胞を持っていたと思われるが、2億2500万年前には、最初の真の哺乳類と言われるアデロバシレウスが出現した。これら初期の哺乳類は(恐竜などの爬虫類との競争を避けたことで)主に夜行性であったため、色覚は生存に必須ではなかった。結果、4タイプのうち2タイプの錐体細胞を失い、青を中心に感知するS錐体と赤を中心に感知するL錐体の2錐体のみを保有するに至った。これは赤と緑を十分に区別できないいわゆる「赤緑色盲」の状態である。この色覚が哺乳類の子孫に遺伝的に受け継がれることとなった。
ヒトを含む旧世界の霊長類(狭鼻猿類)の祖先は、約3000万年前、X染色体にL錐体から変異した緑を中心に感知する新たなタイプの錐体(M錐体)視物質の遺伝子が出現し、ヘテロ接合体の2本のX染色体を持つメスのみが3色型色覚を有するようになり、さらにヘテロ接合体のメスにおいて相同組換えによる遺伝子重複の変異を起こして同一のX染色体上に2タイプの錐体視物質の遺伝子が保持されることとなりX染色体を1本しか持たないオスも3色型色覚を有するようになった。これによって、第3の錐体細胞が「再生」された。3色型色覚はビタミンCを豊富に含む色鮮やかな果実等の発見に有利だったと考えられる。
時代を下ってヒトの色覚の研究成果により、ヒトが属する狭鼻猿類のマカク類に色盲がヒトよりも非常に少ないことを考慮すると、ヒトの祖先が狩猟生活をするようになり3色型色覚の優位性が低くなり、2色型色覚の淘汰圧が下がったと考えられる。色盲の出現頻度は狭鼻猿類のカニクイザルで0.4%、チンパンジーで1.7%である。広鼻猿類でもヨザルは1色型色覚であり、ホエザルは狭鼻猿類と同様に3色型色覚を再獲得している とされている。他方、ホエザルは一様な3色型色覚ではなく、高度な色覚多型であるとの指摘もある。これらのヨザル、ホエザルを除き残りの新世界ザル(広鼻下目)はヘテロ接合体のX染色体を2本持つメスのみが3色型色覚を有し、オスは全て色盲である。これは狭鼻下目のようなX染色体上での相同組換えによる遺伝子重複の変異を起こさなかったためである。ヒトは上記のような初期の哺乳類と霊長目狭鼻下目の祖先のX染色体の遺伝子変異を受け継いでいるため、L錐体のみを保持したX染色体に関連する赤緑色盲が伴性劣性遺伝をする。男性ではX染色体の赤緑色盲の遺伝子を受け継いでいると色盲が発現し、女性では2本のX染色体とも赤緑色盲の遺伝子を受け継いでいる場合に赤緑色盲が発現する。なお、日本人では男性の4.50%、女性の0.165%が先天赤緑色覚異常で、白人男性では約8%が先天赤緑色覚異常であるとされる(詳細は「色覚異常」を参照のこと。)。
狭鼻猿類と広鼻猿類の分岐からさらに時代を下って、ヒト上科とオナガザル上科が分岐したのは、2,800万年から2,400万年前頃であると推定されている。この際に、尿酸オキシダーゼ活性が消失したものと推定される。尿酸オキシダーゼ活性の消失の意味付けは、尿酸が抗酸化物質として部分的にビタミンCの代用となるためである。しかし、ヒトを含むヒト上科では、尿酸オキシダーゼ活性の消失により難溶性物質である尿酸をより無害なアラントインに分解できなくなっているため尿酸が体内で析出する痛風に罹患することがある。
その後の人類への進化については人類の進化を参照のこと。
人間との関係
日本の霊長類研究
第二次世界大戦後、今西錦司らが宮崎県の幸島(こうじま)および高崎山で野生ニホンザル群の餌付けに成功して以来、日本の霊長類研究は飛躍的な発展を遂げた。今西らのニホンザルの文化的行動についての研究は世界中から注目され、その後の霊長類研究の方向性に重大な指針を与えた。
その後もニホンザルにとどまらず、伊谷純一郎など多くの日本人が、ゴリラ、チンパンジー、ボノボなどの類人猿をはじめ、東南アジアからインドにかけてのオナガザル、南米における新世界ザルなど、ほとんどすべてのサルを網羅したフィールドワークを行い、先導的な研究を続けている。
日本のサル学は生態学的研究だけでなく、社会学、生理学、遺伝学、形態学、運動学など多岐に渡り、主に京都大学や大阪大学において今日も活発な研究がなされている。