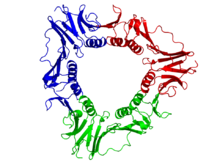
ヒトPCNAスライディングクランプ(虹色、N末端=青、C末端=赤)の上面図(上)と側面図(下)。中央の細孔に二本鎖DNA(マゼンタ)が貫通している。

PolD-PCNA複合体によるDNA結合の構造的基盤
DNAクランプ(DNA clamp)又はスライディングクランプ(sliding clamp)とは、DNA複製を進行させる機能を持つタンパク質複合体である。DNAポリメラーゼホロ酵素の重要な構成要素の1つとしてクランプタンパク質はDNAポリメラーゼに結合し、このDNAポリメラーゼがDNA合成のために鋳型DNAと結合している際に鋳型鎖から解離することを防ぐ。クランプ-ポリメラーゼタンパク質間相互作用は、ポリメラーゼと鋳型DNA鎖の間の直接的な相互作用よりも強力かつ特異的である。ポリメラーゼとDNA鎖の結合はDNA合成反応の律速段階の一つであるため、DNAクランプにより解離と再結合の必要性が無くなることで、DNAクランプが存在しない場合と比較して最大1,000倍ほどDNA合成速度が上がる。
構造
DNAクランプのフォールディング構造はα+βタンパク質であり、ポリメラーゼが伸長中の鎖にヌクレオチドを追加すると、DNA二重らせんを完全に取り囲む多量体構造に組み立てられる。このリング形状により、DNAクランプ及びそれに強力かつ特異的に結合するDNAポリメラーゼはDNA鎖から解離しなくなる。DNAクランプ多量体は複製フォーク上で組み立てられ、ポリメラーゼの前進に伴ってDNA上を「スライド」する。この移動は、クランプタンパク質の中央の穴と、そこを貫通するDNAとの間にある水分子の層によって補助される。
DNAクランプは、真正細菌、古細菌、真核生物、及びいくつかのウイルスにみられる。細菌DNAクランプはDNAポリメラーゼIIIのβサブユニット2個で構成されるホモ二量体であるため、βクランプ(beta clamp)と呼ばれる。古細菌と真核生物ではホモ三量体であり、増殖細胞核抗原(PCNA)と呼ばれる。 T4バクテリオファージは、gp45と呼ばれるDNAクランプも利用する。gp45は、PCNAと構造が似ているが、PCNAまたはβクランプと配列相同性がない三量体である。
|
界
|
DNAクランプタンパク質
|
多量体
|
結合するDNAポリメラーゼ
|
|
真正細菌
|
polIIIのベータサブユニット
|
二量体
|
DNAポリメラーゼIII
|
|
古細菌
|
古細菌PCNA
|
三量体
|
polε
|
|
真核生物
|
PCNA
|
三量体
|
DNAポリメラーゼδ
|
|
ウイルス
|
gp43 / gp45
|
三量体
|
RB69 Pol / T4 Pol
|
細菌DNAクランプ
細菌DNAクランプはDNAポリメラーゼIIIホロ酵素のβサブユニットの二量体であり、βクランプ(beta clamp)と呼ばれる。DNA複製の際にDNAポリメラーゼIIIホロ酵素のγサブユニットがATP加水分解によってエネルギーを得て、2個のβサブユニットからβクランプの組み立てを触媒する。βサブユニットは3つのトポロジー的に同等のドメイン(N末端、中央、C末端)で構成されており、2個のβサブユニットが緊密に会合し、二本鎖DNAを取り囲む閉環を形成する。このDNAとβクランプの結合体は、複製前複合体と呼ばれる。この組み立ての後、βクランプはγサブユニットからαサブユニットとεサブユニットに対して特異的な親和性で結合し、これらが組み合わさって完全なDNAポリメラーゼIIIホロ酵素が形成される。
βクランプは二本鎖DNA上をスライドして移動し、その次にαεポリメラーゼ複合体に結合する。αサブユニットはDNAポリメラーゼ活性を持ち、εサブユニットは3'-5'エキソヌクレアーゼである。
| DNAポリメラーゼIIIのβサブユニット |
|---|
| 識別子 |
|---|
| 略号 |
DNA_polIII_beta |
| Pfam |
PF00712
|
| InterPro |
IPR001001
|
| SMART |
SM00480
|
| SCOP |
2pol
|
| SUPERFAMILY |
2pol
|
| 利用可能な蛋白質構造: |
|---|
| Pfam |
structures
|
| PDB |
RCSB PDB; PDBe; PDBj
|
| PDBsum |
structure summary
|
| PDB |
1jqj, 1jql, 1mmi, 1ok7, 1unn, 1vpk, 2pol, 3bep, 3d1e, 3d1f, 3d1g |
|
|
特定の非ステロイド性抗炎症薬(カプロフェン、ブロムフェナク、及びベダプロフェン)は、細菌のDNAクランプを阻害することにより細菌のDNA複製をある程度抑制する。
真核生物DNAクランプ
真核生物のDNAクランプは、増殖細胞核抗原(PCNA: proliferating cell nuclear antigen)と呼ばれるDNAポリメラーゼδの特定サブユニット3つから組み立てられる。サブユニットのN末端ドメインとC末端ドメインはトポロジー的に同一であり、また3つのサブユニットは緊密に結合し、中心の孔に二本鎖DNAを通した閉環を形成する。
PCNAの配列は、植物、動物、真菌に至るまで真核生物の間でよく保存されており、真核生物全体で同様のDNA複製メカニズムが保存されていることを暗示する。PCNAのホモログは、古細菌(ユーリ古細菌門とクレン古細菌門)及び核多角体病ウイルスにおけるPBCV-1で見出されている。
ウイルスDNAクランプ
ウイルスのgp45スライディングクランプは三量体であり、そのサブユニットタンパク質には2つのドメインがあり、各ドメインは、2つのαヘリックスと2つのβシートで構成されている。フォールドは重複しており、内部に擬2回対称性がみられる。3個のgp45分子が密接に関連して、二本鎖DNAを取り囲む閉環を形成する。
クランプローダー
DNAクランプは、クランプローダー(clamp loader)という特殊なタンパク質によって鋳型鎖にロードされ、DNA複製が完了した後に分解される。DNAクランプ上のこれらの複製開始タンパク質のための結合部位はDNAポリメラーゼのための結合部位と重複しているため、DNAクランプはクランプローダーとDNAポリメラーゼに同時に結合することはできない。したがって、ポリメラーゼと結合している間は、クランプは分解されない。DNAクランプは、ヌクレオソーム会合因子、岡崎フラグメントリガーゼ、 DNA修復タンパク質など、DNA及びゲノムの恒常性に関与する他の因子とも結合する。これらのタンパク質全てにおいて、DNAクランプ上の結合部位はクランプローダーのためのものと共通しているため、これらの酵素がクランプと結合することによって安定的にDNAに作用している間はクランプは取り外されず、また酵素も機能し続ける。クランプローダーがDNAを取り囲むクランプを「閉じる」ためには、ATPの加水分解によるエネルギーが必要となる。
脚注
参考文献
外部リンク