
ミトコンドリア(英語: mitochondrion、複数形: mitochondria)は、ほとんど全ての真核生物の細胞の中に存在する、細胞小器官の1つである。ヤヌスグリーンによって青緑色に染色される。

| 細胞生物学 | |
|---|---|
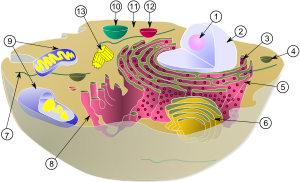
| 動物細胞の模式図 | |
|
ミトコンドリアは脂質二重層でできた外膜と内膜を有し、膜には様々なタンパク質が存在する。ミトコンドリアでは、高エネルギーの電子と酸素分子を利用して、ATPを合成する。すなわち、ミトコンドリアは真核生物における好気呼吸の場である。また、真核生物の細胞が有する核とは別に、ミトコンドリア独自のミトコンドリアDNA(mtDNA)を内部に有し、ある程度ながら自立的にミトコンドリアは細胞内で分裂して、増殖する。このmtDNAは、ミトコンドリア内部だけに限らず、真核生物の細胞全体の生命現象にも関与する。さらに、細胞のアポトーシスにおいても、ミトコンドリアは重要な役割を担っている。
ヒトにおいては、肝臓、腎臓、筋肉、脳などの代謝の活発な細胞には特に多くのミトコンドリアが存在し、細胞質の約40パーセントを占めている。全身の平均では、1細胞中に300個から400個のミトコンドリアが存在し、全身で体重の約1割を占めていると概算されている。単語の「Mitochondrion」はギリシャ語のμίτος, mitos「糸」とχονδρίον, chondrion, 「顆粒」に由来し、糸粒体(しりゅうたい)と和訳される例も見られる。
構造

ミトコンドリアの直径は0.5 μm程度であるが、その形状は、生物種や細胞の置かれている条件によって多様である。球形、円筒形、紐状、網目状など様々な形状のミトコンドリアが存在し、長さが10 μmに達する物も珍しくない。1細胞あたりの数は、1つに維持されている細胞も有るが、多い場合では数千個のミトコンドリアが絶えず分裂と融合を繰り返している場合もある。
ミトコンドリアは外膜と呼ばれる脂質膜に囲まれており、その内側に、もう1枚、内膜と呼ばれる脂質膜を有する。内膜に囲まれた内側をマトリクス、内膜と外膜に挟まれた空間を膜間腔と呼ぶ。なお、内膜はマトリクスに向かって陥入した、クリステ(cristae)と呼ばれる特徴的な構造を取っている。参考までに、この「cristae」とは「櫛」という意味である。
ミトコンドリアは照射された光を強く屈折するため、生きた細胞を位相差顕微鏡で観察すると、ミトコンドリアが明瞭に確認できる。生きた細胞を観察すると、ミトコンドリアが細胞内で、伸縮したり、屈曲したりと、動いている姿も確認できる。
外膜
真核生物の細胞膜と同様に、ミトコンドリアの外膜の組成も、タンパク質とリン脂質の重量比が約1:1である。外膜の進化的起源は真核生物の細胞内膜系だと考えられ、現在でも小胞体膜と物理的に関係しており、カルシウムシグナルの伝達や脂質の交換を行っている。
外膜にはポリンと総称される膜タンパク質が大量に存在し、分子量5000以下の分子が、外膜を通過できるようなチャネルを形成している。これより大きなタンパク質は自由に出入できず、タンパク質のペプチド配列中に、特別な移行シグナルが付与されている場合にのみ、細胞質からミトコンドリア内へと取り込まれる。
膜間腔
膜間腔は、ミトコンドリアの外膜と内膜に挟まれた空間である。外膜がポリンによって低分子を自由に透過させる性質を実現しているため、通常の状態において、膜間腔のイオンや糖などの組成の多くは、ほとんど細胞質と同等である。例外は、内膜の直近のプロトンの濃度のように、限られる。その一方で、膜間腔におけるタンパク質の組成は、細胞質と大きく異なっており、外膜が破壊されて膜間腔に存在するタンパク質(シトクロムcなど)が細胞質へと漏れ出すと、細胞のアポトーシスが引き起こされる。
内膜
内膜はマトリクスと膜間腔とを隔てており、ミトコンドリアの機能的アイデンティティを担っている。酸化的リン酸化に関わる呼吸鎖複合体などの酵素群が、内膜には規則的に配列している。外膜とは対照的に、基本的に内膜は不透性であり、何らかの物質を内膜を横断して輸送するためには、それぞれの物質に対して特異的な輸送体が必要である。
呼吸鎖複合体は内膜を挟んで、マトリクスからプロトンを膜間腔へと汲み出して、膜間腔の側のプロトンの濃度を高め、濃度勾配が形成される。この濃度勾配が、物質輸送やATP合成に関与している。
また、マトリクスへのタンパク質輸送装置やミトコンドリアの分裂・融合に関わるタンパク質群などが存在し、ミトコンドリアを構成する全タンパク質のおよそ2割(150以上)が含まれている。タンパク質とリン脂質の重量比は3:1ほどである。内膜の進化的起源は共生細菌の細胞膜を由来としており、内膜に特徴的なリン脂質カルジオリピンの存在がその証左と考えられている。
一般的に内膜は内側へ向かって陥入し、クリステと呼ばれる構造を形成している。これによって内膜の表面積の増大、ひいてはATP合成能の増大に寄与している。外膜と内膜の表面積の比は細胞のATP需要と相関しており、肝臓では5倍ほど、骨格筋ではさらに大きな値である。
クリステの形状は生物によって様々であり、平板状、管状、団扇状、などが知られている。多細胞動物や陸上植物ではミトコンドリアの長軸に直交する平板状をしており、日本では、教科書などを通じて広く知られている形状である。しかし、これはむしろ特殊な形状であり、真核生物全体を見渡すと、管状のクリステが一般的である。
さらに、同一個体であっても、組織によってクリステの形状が異なる場合がある。例えば、ヒトの多くの細胞のミトコンドリアのクリステは平板状だが、副腎皮質や精巣や卵巣でステロイドホルモン類を分泌する細胞が有するミトコンドリアのクリステは、管状や小胞状であったりする。他にも、ラットでも、このような組織によって、ミトコンドリアのクリステの形状が異なっている事が確認された。さらには、哺乳類のステロイドホルモン分泌細胞以外でも、平板状だけでなく、管状や小胞状のクリステも有するミトコンドリアが観察される場合もある。これらのように例外も数多い。
マトリクス
内膜に囲まれた内側がマトリクスであり、TCAサイクル(別名:TCA回路・クレブス回路・クエン酸回路・クエン酸サイクル)やβ酸化など、ミトコンドリアの代謝機能に関わる酵素群が数多く存在している。ここにはmtDNAが含まれており、ミトコンドリア独自の遺伝情報が保持されている。その遺伝子発現を担うために、リボソーム、tRNA、転写因子や翻訳因子なども存在している。ミトコンドリア全タンパク質の6割から7割が存在しており、非常にタンパク質濃度の高い区画である。
機能
ミトコンドリアの主要な機能は、解糖系やTCAサイクルなどで生成した産物を利用して、電子伝達系に高エネルギーの電子を送り込み、それを酸素に押し付けながら作り出したプロトンの濃度勾配で、ATP合成酵素を駆動して、ADPを酸化的リン酸化によってATPに変換する機能である。。
もちろん、ミトコンドリアが関与しない解糖系のようなATP産生系も存在するものの、真核生物の細胞の活動に必要なATPの多くは、直接、あるいは間接的にミトコンドリアからATPの形で供給される。さらに、ミトコンドリアで行われる、TCAサイクル自体でも実質上はATPと等価なGTPも産生されるなどするため、比喩的に「真核細胞のエネルギーを作り出す場」などと説明される場合もある。
ただし、ATPやGTPの合成以外にもミトコンドリアは多様な機能を有している。例えば、ステロイドやヘムの合成などを含む様々な代謝、カルシウムや鉄の細胞内濃度の調節、細胞周期やアポトーシスの調節などにも大きく関わっているとされる。しかし、これらの機能を全てのミトコンドリアが担っている訳ではなく、機能によっては、特定の細胞でのみ動いている。
こうした様々な機能には多数の遺伝子が関わっており、それらに関わる遺伝子の変異が発生した細胞が、自然免疫で排除されないと、ミトコンドリアの機能低下を招き、ミトコンドリア病を引き起こす場合がある。
ATP産生
ATP産生はミトコンドリアの主たる機能であって、これに関わる多くのタンパク質が内膜やマトリクスに存在している。
細胞質では解糖系が行われ、主にグルコースを代謝して、わずかなATPを合成しながら、ピルビン酸とNADHに分解する。ここで、もし酸素が充分に存在しない場合には、解糖系の産物は嫌気呼吸により代謝される。しかしミトコンドリアで酸素を用いて、これらを酸化する好気呼吸を行えば、嫌気呼吸と比べて効率良くATPを得られる。嫌気性分解では1分子のグルコースから2分子のATPしか得られなかったのに対して、ミトコンドリアによる好気性分解によって、1分子のグルコースから約38分子のATPが合成できる。
また、ミトコンドリアでは、ピルビン酸だけでなく、脂肪酸も利用できる。ミトコンドリアで脂肪酸はβ酸化が行われる。ピルビン酸がアセチルCoAに変換されて、TCAサイクルに入るように、β酸化によって、脂肪酸は炭素鎖が2つずつ切り離されてアセチルCoAが生成され、同じようにTCAサイクルに入るからである。
なお、植物のミトコンドリアは、酸素が無くとも、亜硝酸を利用してある程度のATP産生が可能である。

細胞質での解糖系
地球上の全ての生物で解糖系は、その反応が細胞質基質で起こる。これは解糖系が細胞内小器官が発生する以前から存在してきた、最も原始的な代謝系であることを反映しているのだろうと考えられている。
真核生物では、解糖系で得られた物質(ピルビン酸とNADH)を、TCAサイクルや電子伝達系の反応を行うミトコンドリアへ輸送し、好気呼吸を行う。
ミトコンドリアへの輸送
細胞質の解糖系で生成されたピルビン酸は、ピルビン酸共輸送体(ピルビン酸/H+)により細胞質からミトコンドリアへ輸送される。同じく細胞質で生成されたNADHはリンゴ酸-アスパラギン酸シャトルによりミトコンドリアへ実質的に輸送される。ただし、グリセロリン酸シャトルで輸送される場合もあり、この場合にNADHは、ミトコンドリアのTCAサイクルで発生するFADH2相当に、ミトコンドリアでのATPの産生量は目減りする。
なおADPは、ATP/ADPトランスポーターにより細胞質からミトコンドリアへ輸送される。
また、H2O、O2、CO2、NH3は、そのままミトコンドリア内膜を通過できる。
ピルビン酸の脱炭酸
アセチルCoAは、好気性細胞呼吸の第2段階目である、ピルビン酸がピルビン酸デヒドロゲナーゼ複合体により脱炭酸して生成する。この酵素反応はミトコンドリアのマトリクスで起こる。ここで生成したアセチルCoAは、TCAサイクルに投入される。
TCAサイクル
解糖系で生じたピルビン酸は内膜を能動輸送によって透過し、マトリクスで酸化され補酵素Aと結合し、二酸化炭素、アセチルCoA、NADHを生じる。アセチルCoAは、TCAサイクルへ入る基質である。TCAサイクルの反応に関わる酵素群は、ほとんどがミトコンドリアのマトリクスに存在している。しかし、コハク酸デヒドロゲナーゼだけは例外で、内膜の内側に付着しており、これが電子伝達系の複合体IIに当たる。TCAサイクルで、コハク酸からフマル酸に変換する際の酸化還元反応では、電子伝達系の複合体Iを動かす程のエネルギーが無く、複合体IIが動かされる。その後は、いずれも電子伝達系の複合体IIIへとエネルギーが伝達され、ATP産生に寄与する。
なお、TCAサイクルはアセチルCoAを酸化して二酸化炭素を生じ、その過程で3分子のNADHと1分子FADH2、1分子のGTPを生成する。なお、二酸化炭素はミトコンドリア外へ拡散して排出される。
TCAサイクルでは、サイクルの1回転ごとに、全ての中間体(例えば、クエン酸、イソクエン酸、α-ケトグルタル酸、コハク酸、フマル酸、リンゴ酸およびオキサロ酢酸)が再生される。したがって、ミトコンドリアにこれらの中間体のいずれかを追加して加えると、追加された量がTCAサイクル内に保持され、中間体の1つが他方に変換されて順次増加する。よって、それらの中間体のいずれか1つをTCAサイクルに加えれば、補充反応(アナプレロティック反応)効果を示す。逆に、中間体のいずれかの除去すれば、消費反応(カタプレロティック反応)効果を示す。これらの補充反応及び消費反応は、TCAサイクルの回転で、アセチルCoAと結合してクエン酸を形成するために利用可能な、オキサロ酢酸の量を増加または減少させる。この回転量が、ミトコンドリアによるATP製造量と、細胞へのATPの提供量の増減を左右する。要するに、ミトコンドリア内に存在するTCAサイクルの各種中間体の量が、TCAサイクルの反応速度を調節し、ATPの合成量も調節する事を意味する。
電子伝達系
NADHやFADH2が有する還元力は、内膜にある電子伝達系で数段階を経て、最終的に酸素に渡される。要するに、電子を、電気陰性度の高い酸素に押し付ける形である。
なおNADHは、マトリクスでのTCAサイクルやβ酸化だけでなく、細胞質の解糖系でも生ずる。細胞質で生じたNADHの還元力は、マロン酸-アスパラギン酸対向輸送系や、リン酸グリセロールシャトル系を通じて電子伝達系に供給される。内膜の電子伝達系には、NADH脱水素酵素、シトクロームc還元酵素、シトクロームc酸化酵素が存在しており、プロトン(H+)を膜間腔へ汲み出す。この過程は非常に効率的だが、不充分な反応により活性酸素種を生じ得る(活性酸素#活性酸素と人体の関係参照のこと)。これがいわゆる「酸化ストレス」の形態の1つであり、ミトコンドリアの機能低下や老化に関与していると考えられている。
グルコーストランスポーターであるGLUT1を介して、デヒドロアスコルビン酸がミトコンドリアに輸送され、その後アスコルビン酸に還元され、活性酸素によるフリーラジカルの大部分が生成される場所であるミトコンドリアに蓄積される。アスコルビン酸は、ミトコンドリアの脂質膜とmtDNAを、活性酸素による酸化から保護する。
電子伝達系で、複合体Iと複合体IIIと複合体IVは、電子が伝達された際に、ミトコンドリアのマトリクスから膜間腔へとプロトンを汲み出す。このようにしてプロトンが膜間腔へ汲み出された結果、ミトコンドリアの内膜の隔てて、プロトン濃度の差(電気化学的勾配)が生じる。汲み出されたプロトンは、ATP合成酵素を通じてマトリクスへ戻る事ができ、この際に、電気化学的勾配のポテンシャルを使って、ADPと無機リン酸(Pi)を、ATPへと変換する。生成されたATPは、ATP/ADPトランスポーターによって、ミトコンドリアから細胞質へ輸送され、細胞の活動エネルギー源として利用される。
この原理を化学浸透説と呼び、これをピーター・ミッチェルが最初に唱えた功績によって、1978年にノーベル化学賞を受賞した。また、ATP合成酵素の反応機構を明らかにしたポール・ボイヤーとジョン・E・ウォーカーには、1997年にノーベル化学賞が授与された。
脂肪酸の輸送とβ酸化
ミトコンドリアにおける脂肪酸の輸送については、β酸化#脂肪酸の動員及びβ酸化#脂肪酸の活性化とミトコンドリア内への輸送を参照のこと。ミトコンドリアにおけるβ酸化については、β酸化#β酸化反応および酵素群を参照のこと。
なお、このミトコンドリアのマトリクスで行われる脂肪酸のβ酸化によって、1分子のアセチルCoAを生成する反応の際に、1分子のATPを消費するものの、FADH2とNADHと1分子ずつ生成する。このFADH2とNADHは、電子伝達系に使用され、より多くのATPを産生できる。さらに、ミトコンドリアのマトリクスで生成されたアセチルCoAは、同じくマトリクスで行われているTCAサイクルに投入され、さらに、GTPやATPを産生できる。
アセトアルデヒドの酸化
ミトコンドリアにはアルデヒドデヒドロゲナーゼも発現している。
飲酒などによってエタノールを体内に摂取すると、肝臓などで発現しているアルコールデヒドロゲナーゼなどの作用によって代謝され、アセトアルデヒドが生成する。このアセトアルデヒドを、ミトコンドリアはアルデヒドデヒドロゲナーゼで代謝して、酢酸に変換できる。このミトコンドリアでのアルデヒドデヒドロゲナーゼの活性が遺伝的に低いヒトが、東洋人などの一部に見られ、そのようなヒトはアセトアルデヒドの毒性が強く出易い。
なお、酢酸とは、炭素鎖2つの脂肪酸である。
筋肉とミトコンドリア
速筋線維はミトコンドリアが少なく、グリコーゲンが比較的多いので白く見える。解糖系でATPを産生し、その結果として蓄積したピルビン酸は、乳酸デヒドロゲナーゼで乳酸へと変換され易い。このような嫌気的な糖分解によるATP産生であれば、わざわざ外部から酸素を取り込む必要も無く、速くATPを作り出せる。この事もあり、乳酸性閾値よりも高い運動強度では、速筋線維が多く使われるようになる。しかしながら、この方法では長時間の運動は続けられないという欠点が有る。
これに対して、遅筋線維や心筋は、ミオグロビンが多いので赤く見え、酸素を利用し易い環境を備えている。赤色の筋肉では、乳酸を作るよりは、解糖系の産物であるピルビン酸をミトコンドリアのTCAサイクルへ、解糖系で生成したNADHもミトコンドリアに渡され、ATPを合成して、運動のために使っている。この方式であれば、乳酸などが蓄積しないので、運動強度が低い場合は遅筋線維が主として働いている。
なお、速筋線維で発生した乳酸は、血液を介して肝臓に運ばれ、コリ回路でATPを消費してグルコースの再生に使われる事は、よく知られている。
これ以外に、乳酸デヒドロゲナーゼは、乳酸をピルビン酸に戻す逆反応も触媒できる。遅筋線維や心筋では、外部から取り込んだ乳酸を、ピルビン酸に戻して、ミトコンドリアのTCAサイクルに投入する事も行っている。
いずれにしても、主に速筋線維で蓄積し易い乳酸の代謝には、細胞膜を通過して他の細胞へと乳酸が輸送される必要がある。この乳酸の輸送は、乳酸だけでなくピルビン酸などの輸送にも関わるため、モノカルボン酸の輸送担体(英語: Monocarboxylate Transporter (MCT))と呼ばれている。
熱産生
ある条件下では、膜間腔のプロトンはATP合成に関与せずに、促進拡散によってマトリクスに戻る場合がある。これは「プロトンのリーク」とか「ミトコンドリアの脱共役」と呼ばれ、これによって蓄積されていた電気化学ポテンシャルは熱として解放される。
サーモジェニンなどの一群のプロトンチャネル(脱共役タンパク質)が媒介しており、筋肉の震えを伴わない熱産生に関わっている。サーモジェニンは、若齢や冬眠中の哺乳類に見られる褐色脂肪組織のミトコンドリアに多く存在している。
アポトーシス

細胞に発生したDNA損傷などのストレスは、アポトーシス誘導分子p53やアポトーシスを調節するBcl-2ファミリータンパク質を介して、ミトコンドリアの膜電位を変化させ、外膜の電位依存性陰イオンチャネルが閉鎖される。なお、ミトコンドリアの外膜の電位依存性陰イオンチャネルが閉鎖されると、ミトコンドリアの機能は低下する。
さらに、ミトコンドリアの膜電位の変化は、ミトコンドリアからのシトクロムcの漏出も発生させ、アポトーシスへとつながる。シトクロムcは、細胞質に存在するApaf-1やカスパーゼ-9と結合して、アポトソーム(英語: apoptosome)と呼ばれる集合体を形成する。これによって活性化されたカスパーゼ-9が、下流のエフェクターを活性化する。この後は、DNAが切断されて、細胞は自殺する。
カルシウム貯蔵

細胞内のカルシウム濃度は様々な機構によって厳密に制御されており、細胞中の情報伝達に重要な役割を果たしている。細胞内のカルシウム濃度の上昇により、セカンドメッセンジャー系が起動されたり、筋肉の収縮が起きたりと、様々な反応が起きる。細胞内におけるカルシウムの貯蔵場所としては小胞体が最も顕著だが、カルシウムの貯蔵に関して、小胞体とミトコンドリアは協調している。
と言うのも、ミトコンドリアは一過的なカルシウム貯蔵能を有し、細胞におけるカルシウム濃度の恒常性に貢献しているのである。ミトコンドリアは迅速にカルシウムを取り込む事が可能で、カルシウムは内膜のカルシウム輸送体により、マトリクスへと取り込まれる。これの動作は、ミトコンドリアの膜電位に依存している。
こうして取り込んだカルシウムを、ミトコンドリアが後々放出する事で、カルシウム濃度の緩衝作用を果たしている。なお、カルシウムの放出は、ナトリウム・カルシウム対向輸送、もしくは、カルシウム依存性カルシウム放出系によって行われる。
ミトコンドリアゲノム
ミトコンドリア中には、細胞核とは別に、独自のDNAが存在しており、これをミトコンドリアDNA(mtDNA)と呼ぶ。mtDNAは、細胞核とは異なる独自の遺伝情報を持っている。DNA分子の大きさや形状、codeされている遺伝子の数や種類などは、生物種によって大きく異なる。
ただ、通常はGC含量が低く(20-40%)、基本的なmtDNAは、塩基対が数十kb程度のDNAである。mtDNAには、電子伝達系に関わるタンパク質、リボソームRNAやtRNAなど、数十種類の遺伝子がcodeされている。
ヒトを含む脊椎動物のmtDNAは、真核生物の中ではかなり特殊な性質を多く持っており、研究はよく進んでいるものの、安易な一般化は慎まなければならない。
なお、mtDNAと、それに基いて合成される産物の一部は、ミトコンドリアだけではなく、細胞表面にも所在し、mtDNAに突然変異が発生している場合には、自然免疫系が特異的に細胞ごと破壊して排除する。mtDNAに突然変異が発生した場合には、ミトコンドリア病を発症する可能性もある。
mtDNAの塩基対数と形状
最も小さなmtDNAを持つ生物はアピコンプレックス門の原虫で、大きさわずか6 kbの線状ゲノムである。電子伝達系に関わる3つのタンパク質遺伝子と、断片化されたリボソームRNA遺伝子群のみが存在している。逆に最も大きなmtDNAは、マスクメロンの持つ2400 kbという巨大なゲノムである。ただし遺伝子数は比較的多いものの、それでも100弱に過ぎず、大量の反復配列やグループ2イントロンなどの非遺伝子領域が大部分を占める。
ヒトを含む多細胞動物のmtDNAはいずれも比較的似通っており、長さ16 kb前後の単一の環状DNAで構成されている。遺伝子は37あり、その内訳は、呼吸鎖複合体とATP合成酵素のサブユニットが13、tRNAが22、rRNAが2である。
遺伝子地図などでは、mtDNAが環状に表現される事例が多い。しかし物理的に環状のmtDNAを持つ生物はごく一部に限られ、多くの生物では環状の基本構造からトイレットペーパーを引き出すかのように連続的に複製されており、その結果mtDNAの大部分は、基本単位が何度も繰り返す線状反復構造を有している。また少数派ではあるものの、常に線状のmtDNAを持つ生物も存在している。
遺伝子
ミトコンドリアゲノムはαプロテオバクテリアから受け継がれており、その遺伝子発現は細菌と共通した特徴を持っており、真核生物の細胞核のDNAとは異なる。例えば、複数の遺伝子がまとめて転写され、それが遺伝子ごとに切断されポリアデニル化されて成熟mRNAとなる点や、翻訳の開始にフォルミル化メチオニンが利用される点、細胞核に存在するようなスプライソソーム型のイントロンが存在しない点、などが挙げられる。
さらに、ミトコンドリアの遺伝暗号表は、細胞核や一般の原核生物で利用されている普遍暗号表と比べて、若干の差が見られる。顕著な例として、細胞核では終止コドンであるはずのUGAが、ミトコンドリアではトリプトファンをコードしている場合が多い事が挙げられるものの、例外も多く、生物種によって少しずつ異なる暗号表を用いているのが実態である。
またミトコンドリアでは、しばしばRNA編集が行われる。例えば高等植物のミトコンドリアでは、DNA配列上のCGGがmRNAの場合は、UGGと編集されてトリプトファンをコードするという例が知られている。
ただ、重要な点として、ミトコンドリアの機能に関わる全ての遺伝子が、mtDNAに存在しているわけではないが挙げられる。ミトコンドリアが持つmtDNA上にcodeされているミトコンドリアゲノムは、細菌のゲノムと比べると、遺伝子数が極端に減少している。一方で、ミトコンドリアが必要とする大多数の遺伝子は、細胞核の側にcodeされており、細胞質の側で転写された情報に基いて生合成された遺伝子産物が、ミトコンドリアへと輸送される。これは進化の過程で、遺伝子が細胞核へ移動したからだと考えられている。
こうした現象は、比較的よく起きた出来事だと考えられ、マイトソームなどのように全てのDNAを完全に失ったようなミトコンドリアも存在している。
一方で、原生生物のレクリノモナス(Reclinomonas americana)は、他の生物では細胞核から輸送されているようなタンパク質の遺伝子が、mtDNA上に存在しており、比較的原始的なミトコンドリアゲノムを未だに保持していると考えられている。
ミトコンドリアには、呼吸機能に関与する疎水性のタンパク質が存在し、疎水性であるために輸送が難しく、これらをミトコンドリアの内部で作らざるを得ないために、ミトコンドリアに遺伝子が残っている理由の1つと考えられている。
異数性
1つのミトコンドリアには、2-10コピーのDNA分子が存在する。その全てが完全に同じ情報を持つわけではなく、複数の異質のDNA分子を含んでいると確認されている。
起源
ミトコンドリアは、リケッチアに近い好気性細菌のαプロテオバクテリアが、真核細胞に入り込んだ結果として獲得されたと考えられている。リン・マーギュリスの細胞内共生説では単に好気性バクテリアが起源とされていたが、その後すぐの1970年代にはすでにミトコンドリアの起源が現在でいうαプロテオバクテリアだという意見が出た。脱窒細菌のParacoccus denitrificansや、暗所好気条件で培養した紅色光合成細菌のRhodobacter sphaeroidesは、呼吸鎖の構成や阻害剤への応答がミトコンドリアと類似しており、特に、シトクロムcがミトコンドリアと互換性を持つ点が注目された。
細胞核DNAにコードされているシトクロムcだけでなく、mtDNAにコードされているリボソームRNAの配列を使った系統解析でもαプロテオバクテリア起源であると示され、1980年代にはミトコンドリアのαプロテオバクテリア起源は受け入れられるようになった。
ただし、初期の解析では高等植物ミトコンドリアのリボソームRNAの配列が、他のミトコンドリアの配列と比べて進化的距離が非常に小さかったため、ミトコンドリアの起源は単独ではなく、高等植物のミトコンドリアは新たに獲得された物だという意見も有った。しかし、こうした意見は現在では否定され、真核生物のミトコンドリアの起源は単一であるとされている。
もっとも、αプロテオバクテリアは非常に多様な細菌を含む分類群であり、その中でどのような細菌がミトコンドリアの起源なのかについては、長く議論が続いている。初期には前述の通り脱窒細菌や光合成細菌が起源だと考えられていたが、シャペロニンHsp60(GroEL)を用いた系統解析によりリケッチアが最も近縁であると示されてからは、これが有力説となった。リケッチアは、細胞内寄生生物である点、TCAサイクルを持ち好気呼吸ができるのに対して解糖系を持たない点、細胞膜にADP/ATP輸送体を持っている点、ゲノムが小さくAT含量が高い点など、ミトコンドリアと共通した特徴が複数見られる。
1998年に発疹チフスを引き起こすリケッチアの1種であるRickettsia prowazekiiのゲノムが解読され、祖先的とされるReclinomonas americanaのミトコンドリアゲノムと共通している遺伝子や、配置順が保存された遺伝子群などが見出され解析された。その多くはミトコンドリアがリケッチアに近縁であるという仮説を支持する結果であったが、ADP/ATP輸送体については、予想に反して起源を異にしていると示された。
20世紀末から21世紀初頭にかけて、世界中の海洋には自由生活性で浮遊性の細菌ペラジバクター(暫定的にCandidatus Pelagibacter ubiqueと命名されている)が存在していることが明らかとなった。ペラジバクターはリケッチア目の中で、最も祖先的な位置から派生したと考えられる生物であり、ミトコンドリアの起源を、ペラジバクターとその他一般的なリケッチアとの間に求められる。
なお、アメーバに似た原生生物であるペロミクサや微胞子虫など、原生生物の中はミトコンドリアを持っていないものもいる。これを、ミトコンドリアが共生する以前の真核生物の生き残りと見る説があった(→アーケゾア仮説)が、後に否定された。
生物の系統との関係
ミトコンドリアの特徴は、動物、植物、菌類にほぼ共通であるが、それ以外の原生動物では、若干異なった形状の物が有る。特にクリステの形については、明らかに異なった形状のミトコンドリアが見られる。
ヒトなどの一般のミトコンドリアでは、内膜がひだのように折れ曲がり、クリステは平坦な板のような形をしている。しかし、粘菌類の場合、クリステは内膜から内部へと放射状に入り込む管の形で、管の表面にATP合成酵素の手段が並んでいる。また、内部の中央にDNAを含んだ塊があって、ミトコンドリア核と呼ばれる。このような、管状のクリステを持つミトコンドリアは、繊毛虫やアピコンプレックス類、アメーバ類、クロララクニオン藻類などの原生生物にも見られる。
また、ミドリムシ類とトリパノソーマでは、クリステは団扇型である。これらのミトコンドリアは、細長くて枝分かれをして、細胞内に広がっている。トリパノソーマでは、鞭毛の基部にキネトプラストと呼ばれる袋状の構造が知られており、その中の顆粒にはDNAが含まれているが、これはミトコンドリアの一部である。
がんとミトコンドリア
1955年にオットー・ワールブルクは、体細胞が長期間低酸素状態に晒されると呼吸障害を引き起こし、通常の酸素濃度の環境下に戻しても、大半の細胞が変性や壊死を起こし、ごく一部の細胞が酸素呼吸に代わるエネルギー生成経路を昂進させて生存する細胞が、ガン細胞になるとの説を発表した。この説では、酸素呼吸よりも、むしろ解糖系によるエネルギー産生に依存する細胞は、下等動物や胎生期の未熟な細胞が一般的であり、体細胞がATP産生を酸素呼吸によらず解糖系に依存した結果、細胞が退化してガン細胞が発生するとした。
ガン細胞の発生とmtDNAの突然変異の関与は、古くから指摘されてきた。その理由は特定の発ガン性化学物質が、DNAよりもmtDNAに結合し易い事と、ガン組織のmtDNAは正常組織よりも高い割合で突然変異の蓄積が観察された事による。しかしながら、母性遺伝するガンの存在が確認されていない点や、DNAの影響を排除しmtDNA単独でのガンへの影響を検証する手法が確立されていない点などが、この仮説の証明の障害であった。
ただ、2008年筑波大学の林純一らが、ガンの転移能獲得という、ガン細胞の悪性化に、mtDNAの変異が関与している事を指摘した。マウス肺がん細胞の細胞質移植による細胞雑種の比較により、mtDNAの特殊な病原性突然変異によってガン細胞の転移能獲得の原因になる事を発見し、ヒトのガン細胞株でも、mtDNAの突然変異がガン細胞の転移能を誘導し得る事を明らかにし、少なくとも、mtDNAがATP合成以外の生命現象にも関与する事を明らかにした。また、林らによるとmtDNAの突然変異には、活性酸素種(ROS)の介在が重要であり、ROSを除去すれば転移能の抑制が可能ではないかと推察した。ただし、ガン細胞の転移能の獲得メカニズムは複雑であり、様々な要因が考えられるので、これはその要因の1つに過ぎない。
また、京都大学の井垣達吏らは、① Ras遺伝子の活性化とミトコンドリアの機能障害を起こした細胞は、細胞老化を起こして細胞老化関連分泌因子(SASP因子)を放出し、これにより周辺組織のガン化を促進する事、また、② 細胞分裂停止とJNK遺伝子の活性化が互いに増幅し合う事で、細胞内のJNK活性が顕著に増大し、これによりSASP因子の産生が誘導される事を示した。
「ミトコンドリア・イブ」
ミトコンドリアのDNAは、同種交配の場合卵子に入った精子のミトコンドリアが選択的に排除されるため、母親のmtDNAを引き継ぐ事を根拠に、現生人類の起源の地が探られた。すなわち、世界中に分布するヒトからmtDNAを調べて、現在の分布地図から現生人類の起源とその移動について推察する作業を実施した。この結果、大昔のアフリカのある女性が、今の人類の全てのミトコンドリアについての「母親」であるとの仮説が発表された。この女性はキリスト教徒の宗教的説話になぞらえて「ミトコンドリア・イブ」と呼ばれている。
しかしながら、この仮説は、その他の遺伝情報について、この女性に全てが由来するという意味ではない。無論、全人類の起源が1人の女性にあると言っているわけでもない。しかも、実験的に異種交配させた受精卵では、精子由来のミトコンドリアを排除するプロセスが失敗する場合がある事が知られている。
フィクション
小説
1995年に第2回日本ホラー小説大賞を受賞した瀬名秀明の『パラサイト・イヴ』は、ミトコンドリアの共生起源説、および、人類の進化におけるミトコンドリア・イブ説に基づき、現在のミトコンドリアは細胞の支配下にあるが、もしもそれが反乱を起こしたならば、という仮定の物語で話題を呼び、映画やゲーム化も行なわれた。
脚注
注釈
関連項目
- ミトコンドリアのシャトル系
- 葉緑体 - ミトコンドリアと同様に、外膜と内膜、さらに、独自のDNAも有する。
- カルニチン
- ハプログループF (mtDNA)
- 母の呪い
- オンコサイト