

電気化学的勾配(でんきかがくてきこうばい、英: electrochemical gradient)とは電気化学ポテンシャルの勾配であり、通常は膜を越えて移動するイオンについてのものである。勾配は、化学的勾配(膜を挟んだ溶質濃度の差)と電気的勾配(膜を挟んだ電荷の差)という2つの部分から構成される。透過性の膜を挟んだ両側のイオン濃度が不均等であるときには、イオンは高濃度側から低濃度側へ単純拡散によって膜を越えて移動する。イオンは電荷を持っているため、膜を挟んで電位も形成される。膜を挟んで電荷が不均等に分布している場合、膜の両側で電荷が均等となるまでイオンの拡散を駆動する力が電位差によって生み出される。
定義
電気化学的勾配は電気化学ポテンシャルの勾配である。






概要
電気化学ポテンシャルは電気分析化学や、バッテリーや燃料電池などの産業的応用に重要である。電気化学ポテンシャルはポテンシャルエネルギーの多くの互換可能な形態のうちの1つである。
生物学的過程では、イオンが膜を越えて拡散または能動輸送される方向は電気化学勾配によって決定される。ミトコンドリアと葉緑体では、プロトン駆動力(proton motive force)と呼ばれる化学浸透ポテンシャルを形成するためにプロトン勾配が利用される。このポテンシャルエネルギーは、酸化的リン酸化や光リン酸化によるATP合成のために利用される。
電気化学的勾配は2つの要素から構成される。1つ目は電気的要素であり、脂質膜を挟んだ電荷の差によって生じる。2つ目は化学的要素であり、膜を挟んだイオン濃度の差によって生じる。これら2つの因子の組み合わせによって、膜を越えたイオンの移動の熱力学的に有利な方向が決定される。
電気化学的勾配は水力発電ダムの水圧に例えられる。膜内部のナトリウム-カリウムポンプなどの膜輸送体は、水のポテンシャルエネルギーを他の物理的・化学的エネルギーに変換するタービンに相当し、膜を通過するイオンはダムの下流に落ちる水に相当する。また、エネルギーを使って水を上流のダム湖へと汲み上げることができるのと同様に、細胞内の化学エネルギーを用いて電気化学的勾配を形成することができる。
生物学
イオンが細胞膜を通過することで形成される膜電位は、神経伝導、筋収縮、ホルモンの分泌、感覚などの生物学的過程を駆動する。典型的な動物細胞では、細胞内の電位は細胞外に対して-50 mVから-70 mVである。
電気化学的勾配は、ミトコンドリアでの酸化的リン酸化におけるプロトン勾配の形成にも関与している。細胞呼吸の最終段階は電子伝達系である。ミトコンドリアの内膜に埋め込まれた4つの複合体が電子伝達系を構成する。しかし、プロトンをマトリックスから膜間腔(IMS)へ汲み上げるのは複合体I、III、IVのみである。系全体として、マトリックスからIMSへ10個のプロトンが移行し、200 mV以上の電気化学ポテンシャルが形成される。これによってプロトンがATP合成酵素を通ってマトリックスへ戻る流れが作られ、ATP合成酵素はADPに無機リン酸を付加することでATPを産生する。このように、プロトンの電気化学的勾配の形成はミトコンドリアでのエネルギー産生に重要である。電子伝達系全体としての反応式は次のようになる。
NADH + 11 H+(matrix) + 1/2 O2 → NAD+ + 10 H+(IMS) + H2O
電子伝達系と同様に、光合成の明反応も葉緑体チラコイドのルーメンへプロトンを汲み上げ、ATP合成酵素によるATPの合成を駆動する。プロトン勾配は循環的光リン酸化または非循環的光リン酸化のいずれかによって形成される。非循環的光リン酸化に関与するタンパク質の中で、光化学系II(PSII)、プラストキノン、シトクロムb6f複合体はプロトン勾配の形成に直接寄与する。4個の光子がPSIIによって吸収され、8個のプロトンがルーメンへ汲み上げられる。光リン酸化全体としての反応式は次のようになる。
2 NADP+ + 6 H+(stroma) + 2 H2O → 2 NADPH + 8 H+(lumen) + O2
他の輸送体やイオンチャネルもプロトンの電気化学的勾配に寄与している。カリウムチャネルTPK3はCa2+によって活性化され、チラコイドルーメンからストロマへK+を透過させることでpH勾配の形成を助ける。一方、電気的に中性なK+排出アンチポーターKEA3はK+をチラコイドルーメンへ、H+をストロ間へ輸送することで電場の形成を助ける。
イオン勾配

イオンは電荷を持つため、単純拡散で膜を通過することはできない。イオンが膜を越えて輸送される機構には、能動輸送と受動輸送の2つの機構が存在する。イオンの能動輸送の例としては、Na+/K+-ATPアーゼ(NKA)が挙げられる。NKAはATPからADPと無機リン酸への加水分解を触媒し、ATP1分子の加水分解ごとに3個のNa+が細胞外へ輸送され、2個のK+が細胞内へ輸送される。その結果、細胞内は細胞外よりも負の電位となり、具体的には約-60 mVの膜電位Vmembraneが生じる。受動輸送の例は、Na+、K+、Ca2+、Cl−チャネルを介したイオンの流れが挙げられる。これらのイオンは濃度勾配に従って移動する。例えば、Na+は細胞外で高濃度であるため、Na+はNa+チャネルを通って細胞内へ流入する。細胞内の電位は負であるため、陽イオンの流入によって膜は脱分極し、膜電位はゼロに近くなる。しかし、化学的勾配の影響が電気的勾配の影響よりも大きい限り、Na+は濃度勾配に従って移動し続ける。双方の勾配の影響が等しくなると(Na+の場合、膜電位が約+70 mVに達すると)、駆動力(ΔG)がゼロとなるためNa+の流入は停止する。駆動力の方程式は次のように表される。

Rは気体定数、Tは絶対温度、zはイオンの電荷、Fはファラデー定数を表している。
細胞内のイオン濃度を次の表に示す。X-はタンパク質の総負電荷を表している。
| イオン | 哺乳類 | イカ軸索 | 出芽酵母 | 大腸菌 | 海水 | ||
|---|---|---|---|---|---|---|---|
| 細胞 | 血液 | 細胞 | 血液 | ||||
| K+ | 100 - 140 | 4-5 | 400 | 10 - 20 | 300 | 30 - 300 | 10 |
| Na+ | 5-15 | 145 | 50 | 440 | 30 | 10 | 500 |
| Mg2+ | 10 0.5 - 0.8 |
1 - 1.5 | 50 | 30 - 100 0.01 - 1 |
50 | ||
| Ca2+ | 10−4 | 2.2 - 2.6 1.3 - 1.5 |
10−4 - 3×10−4 | 10 | 2 | 3 10−4 |
10 |
| Cl− | 4 | 110 | 40 - 150 | 560 | 10 - 200 | 500 | |
| X− | 138 | 9 | 300 - 400 | 5 - 10 | |||
| HCO3− | 12 | 29 | |||||
| pH | 7.1 - 7.3 | 7.35 - 7.45 (動脈血)
6.9 - 7.8 (全血液) |
7.2 - 7.8 | 8.1 - 8.2 | |||
プロトン勾配
プロトン勾配は多くの細胞種でエネルギー貯蔵の形態として重要である。通常、勾配はATP合成酵素、鞭毛の回転、代謝産物の輸送などを駆動するために利用される。この節では各細胞でプロトン勾配の形成を助ける3つの過程、バクテリオロドプシン、非循環的光リン酸化、酸化的リン酸化に焦点を当てる。
バクテリオロドプシン
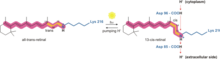
古細菌では、バクテリオロドプシンはプロトンポンプによってプロトン勾配を形成する。プロトン勾配はH+濃度の低い側から高い側へ移動させるプロトン輸送体に依存している。バクテリオロドプシンでは、プロトンポンプは568 nmの波長の光子の吸収によって活性化され、レチナールのシッフ塩基(SB)の異性化によってK状態となる。これによってSBはAsp85とAsp212から離れ、SBからAsp85へのH+の転移が引き起こされてM1状態となる。その後、Glu194からGlu204が引き離されてM2状態に移行し、Glu204から外部溶媒へプロトンが放出される。SBはAsp96によって再プロトン化されN状態となる。Asp96の脱プロトン化状態は不安定であるため、細胞質からのプロトンによって迅速に再プロトン化が行われることは重要である。Asp85とAsp96のプロトン化はSBの再異性化を引き起こしてO状態となる。最終的に、Asp85からGlu204へプロトンが渡されてバクテリオロドプシンは基底状態となる。
光リン酸化
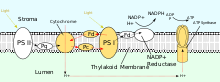
葉緑体におけるPSIIによるプロトン勾配の形成の駆動は光に依存しているが、PSIIはプロトン勾配の形成のために方向性のある酸化還元反応を利用する。タンパク質を介して物理的にプロトンを輸送するのではなく、プロトンの結合を必要とする化学反応が細胞側で、プロトンの遊離を必要とする化学反応が細胞内側で起こることで結果的にプロトンが移行する。まず、P680の2つの電子を高エネルギー状態へ活性化するために、680 nmの波長の光子が吸収される。高エネルギー電子はタンパク質に結合したプラストキノン(PQA)へ移動し、その後非結合状態のプラストキノン(PQB)へ移動する。これによってプラストキノン(PQ)はプラストキノール(PQH2)へと還元され、ストロマから2つの光子を獲得した後PSIIから解離する。P680の電子は酸素発生複合体による水分子の酸化によって補充される。その結果、O2とH+がルーメンへ放出される。全体の反応は次のように表される。

PSIIから解離した後PQH2はシトクロムb6f複合体へ移動し、その後2つの異なる反応で2つの電子がPQH2からプラストシアニンへ移動する。この過程は、電子伝達系の複合体IIIで行われるQサイクルと類似している。最初の反応では、PQH2は複合体のルーメン側に結合し、1つの電子が鉄硫黄中心へ移動し、その後プラストシアニンへ移動する。もう1つの電子はヘムbLへ移動し、その後ヘムbL、プラストキノンへ移動する。2番目の反応では、2つ目のPQH2が酸化され、他のプラストシアニン分子とPQに電子が付加される。双方の反応によって、4つのプロトンがルーメンへ移行する。
酸化的リン酸化
電子伝達系では、複合体Iが還元型ニコチンアミドアデニンジヌクレオチド(NADH)から2つの電子を移動することにより、ユビキノン(UQ)からユビキノール(UQH2)への還元を触媒する。それに伴って、4つのプロトンがミトコンドリアマトリックスからIMSへ移行する。

複合体IIIはQサイクルを触媒する。最初の段階では、複合体Iによって還元されたUQH2からの2つの電子がQo部位の2分子の酸化型シトクロムcへ移動する。2番目の段階では、Qi部位のUQH2をUQへ還元するためもう2つの電子が必要となる。全体の反応は次のように表される。

複合体IVは、複合体IIIで還元されたシトクロムcからの2つの電子の酸素分子の一方への移動を触媒する。酸化的リン酸化で酸素1分子を完全に利用するためには、4つの電子の移動が必要となる。その後、酸素は水分子を形成するためにマトリックスの4つのプロトンを消費し、4つのプロトンがIMSへと汲み出される。全体の反応は次のように表される。

関連文献
- Campbell & Reece (2005). Biology. Pearson Benjamin Cummings. ISBN 978-0-8053-7146-8
- Stephen T. Abedon, "Important words and concepts from Chapter 8, Campbell & Reece, 2002 (1/14/2005)", for Biology 113 at the Ohio State University