
 リガンド依存性イオンチャネル
| |||||||||
| 識別子 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 略号 | Neur_chan_memb | ||||||||
| Pfam | PF02932 | ||||||||
| InterPro | IPR006029 | ||||||||
| PROSITE | PDOC00209 | ||||||||
| SCOP | 1cek | ||||||||
| SUPERFAMILY | 1cek | ||||||||
| TCDB | 1.A.9 | ||||||||
| OPM superfamily | 14 | ||||||||
| OPM protein | 2bg9 | ||||||||
| |||||||||

リガンド依存性イオンチャネル (Ligand-gated ion channels; LIC、LGIC) は、一般的にイオンチャネル型受容体とも呼ばれ、神経伝達物質などの化学的メッセンジャー (すなわちリガンド) の結合に応答して、Na+、K+、Ca2+、Cl-などのイオンが膜を通過するように開く、膜貫通型イオンチャネルタンパク質のグループである。
シナプス前神経細胞が興奮すると、小胞からシナプス間隙に神経伝達物質が放出される。次に、神経伝達物質はシナプス後神経細胞にある受容体に結合する。これらの受容体がリガンド依存性イオンチャネルである場合、結果として生じるコンホメーション変化によりイオンチャネルが開き、細胞膜を横切るイオンの流れが生じる。これにより、興奮性の受容体反応では脱分極 (en:英語版) 、抑制性の受容体反応では過分極が発生する。
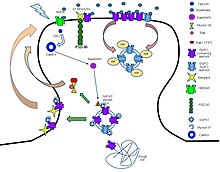
これらの受容体タンパク質は、典型的には、少なくとも2つの異なるドメインから構成されている。イオン孔を含む膜貫通ドメインと、リガンド結合部位 (アロステリック結合部位) を含む細胞外ドメインである。このモジュール性により、タンパク質の構造を見つけるための「分割統治」アプローチが可能になった (各ドメインを別々に結晶化する)。シナプスに位置するこのような受容体の機能は、シナプス前に放出された神経伝達物質の化学信号を直接かつ非常に迅速にシナプス後の電気信号に変換することである。多くのLICは、アロステリックリガンド、チャネルブロッカー、イオン、または膜電位によってさらに調節される。LICは、進化的な関係を持たない3つのスーパーファミリーに分類される。Cysループ型受容体、イオンチャネル型グルタミン酸受容体、ATP依存性チャネルである。
Cysループ型受容体

Cysループ型受容体は、N末端の細胞外ドメインにある2つのシステイン残基間のジスルフィド結合によって形成される特徴的なループにちなんで命名された。これらは通常、このジスルフィド結合を欠いている五量体リガンド依存性イオンチャネルの大規模なファミリーの一部であるため、暫定的な名称である「プロループ受容体」に由来する。細胞外N末端リガンド結合ドメインの結合部位は、脊椎動物では(1)アセチルコリン(AcCh)、(2)セロトニン、(3)グリシン、(4)グルタミン酸、(5)γ-アミノ酪酸(GABA)の受容体特異性を有している。受容体は、それらが伝導するイオンの種類 (アニオン性またはカチオン性) によって細分化され、さらに内因性リガンドによって定義されるファミリーに分類される。それらは通常、五量体であり、各サブユニットは膜貫通ドメインを構成する4回膜貫通ヘリックスを含み、かつβシートサンドイッチ型の細胞外N末端リガンド結合ドメインを含む。また、画像に示すように、細胞内ドメインを含むものもある。
原形のリガンド依存性イオンチャネルはニコチン性アセチルコリン受容体である。これは、タンパク質のサブユニット (通常はααβγδ) の五量体で構成されており、アセチルコリンの2つの結合部位 (各αサブユニットの境界に1つ) がある。アセチルコリンが結合すると、受容体の構成が変化し (T2ヘリックスがねじれ、細孔をブロックするロイシン残基を、チャネル経路の外に移動させる)、細孔の収縮が約3オングストロームから8オングストロームに広がり、イオンが通過できる。この細孔により、Na+イオンが、電気化学的勾配を下って細胞内に流入する。一度に十分な数のチャネルが開くと、Na+イオンによって運ばれた正電荷の内向きの流れが、シナプス後膜を十分に脱分極させて活動電位を引き起こす。
バクテリアのような単細胞生物は、活動電位の伝達をほとんど必要としないが、LICに対するバクテリアのホモログは同定されており、それにもかかわらず化学受容体として作用すると仮定されている。この原核生物の nAChR 変異体は、それが同定された種「Gloeobacter Ligand-gated Ion Channel」にちなんで、GLIC受容体として知られている。
構造
Cysループ型受容体には、αヘリックスと10個のβストランドを持つ大きな細胞外ドメイン (ECD) があり、よく保存された構造要素がある。ECDに続いて、4つの膜貫通セグメント (TMS) が細胞内および細胞外ループ構造によって接続されている。TMS 3-4ループを除いて、それらの長さはわずか7-14残基である。TMS 3-4ループは、細胞内ドメイン (ICD) の最大部分を形成しており、これらの相同受容体の間で最も可変的な領域を示している。ICDは、TMS 3-4ループと、イオンチャネル孔の前のTMS 1-2ループによって定義されている。結晶化により、このファミリーの一部のメンバーの構造が明らかにされているが、結晶化を可能にするために、細胞内ループは通常、原核生物のcysループ型受容体に存在する短いリンカーで置換されているため、その構造は不明である。しかしながら、この細胞内ループは脱感作、薬理学的物質によるチャネル生理機能の調節、および翻訳後修飾で機能しているようである。また、細胞内ループの中には輸送に重要なモチーフが存在し、ICDは抑制性シナプス形成を可能にする足場タンパク質と相互作用する。
カチオン性cysループ型受容体
| Type | Class | IUPHAR-recommended protein name |
Gene | Previous names |
|---|---|---|---|---|
|
Serotonin (5-HT) |
5-HT3 |
5-HT3A 5-HT3B 5-HT3C 5-HT3D 5-HT3E |
HTR3A HTR3B HTR3C HTR3D HTR3E |
5-HT3A 5-HT3B 5-HT3C 5-HT3D 5-HT3E |
|
Nicotinic acetylcholine (nAChR) |
alpha |
α1 α2 α3 α4 α5 α6 α7 α9 α10 |
CHRNA1 CHRNA2 CHRNA3 CHRNA4 CHRNA5 CHRNA6 CHRNA7 CHRNA9 CHRNA10 |
ACHRA, ACHRD, CHRNA, CMS2A, FCCMS, SCCMS |
| beta |
β1 β2 β3 β4 |
CHRNB1 CHRNB2 CHRNB3 CHRNB4 |
CMS2A, SCCMS, ACHRB, CHRNB, CMS1D EFNL3, nAChRB2 |
|
| gamma | γ | CHRNG | ACHRG | |
| delta | δ | CHRND | ACHRD, CMS2A, FCCMS, SCCMS | |
| epsilon | ε | CHRNE | ACHRE, CMS1D, CMS1E, CMS2A, FCCMS, SCCMS | |
|
Zinc-activated ion channel (ZAC) |
ZAC | ZACN | ZAC1, L2m LICZ, LICZ1 |
アニオン性cysループ型受容体
| Type | Class | IUPHAR-recommended protein name |
Gene | Previous names |
|---|---|---|---|---|
| GABAA | alpha |
α1 α2 α3 α4 α5 α6 |
GABRA1 GABRA2 GABRA3 GABRA4 GABRA5 GABRA6 |
EJM, ECA4 |
| beta |
β1 β2 β3 |
GABRB1 GABRB2 GABRB3 |
ECA5 |
|
| gamma |
γ1 γ2 γ3 |
GABRG1 GABRG2 GABRG3 |
CAE2, ECA2, GEFSP3 | |
| delta | δ | GABRD | ||
| epsilon | ε | GABRE | ||
| pi | π | GABRP | ||
| theta | θ | GABRQ | ||
| rho |
ρ1 ρ2 ρ3 |
GABRR1 GABRR2 GABRR3 |
GABAC | |
|
Glycine (GlyR) |
alpha |
α1 α2 α3 α4 |
GLRA1 GLRA2 GLRA3 GLRA4 |
STHE |
| beta | β | GLRB |
イオンチャネル型グルタミン酸受容体
イオンチャネル型グルタミン酸受容体は、神経伝達物質であるグルタミン酸に結合する。それらは、細胞外アミノ末端ドメイン (ATD、四量体の組み立てに関与)、細胞外リガンド結合ドメイン(LBD)、膜貫通ドメイン(TMD)で構成されるサブユニットごとに四量体を形成する。各サブユニットの膜貫通ドメインは、3つの膜貫通ヘリックスと、リエントラントループを持つ半膜ヘリックス(half membrane helix)を含む。タンパク質の構造は、N末端のATDから始まり、続いてLBDの前半がTMDのヘリックス1,2,3によって中断され、その後、LBDの後半に続き、次にC末端のTMDのヘリックス4で終わる。これは、TMDと細胞外ドメインの間に3つのリンクがあることを意味している。四量体の各サブユニットは、クラムシェルのような形状を形成する2つのLBDセクションによって形成されたグルタミン酸の結合部位を持っている。イオンチャネルを開くためには、四量体のこれらのサイトのうちの2つだけが占有される必要がある。細孔は、反転カリウムチャネル(inverted potassium channel)に似た形で、主にハーフヘリックス 2(half helix 2)によって形成される。
| Type | Class | IUPHAR-recommended protein name |
Gene | Previous names |
|---|---|---|---|---|
| AMPA | GluA |
GluA1 GluA2 GluA3 GluA4 |
GRIA1 GRIA2 GRIA3 GRIA4 |
GLUA1, GluR1, GluRA, GluR-A, GluR-K1, HBGR1 GLUA2, GluR2, GluRB, GluR-B, GluR-K2, HBGR2 GLUA3, GluR3, GluRC, GluR-C, GluR-K3 GLUA4, GluR4, GluRD, GluR-D |
| Kainate | GluK |
GluK1 GluK2 GluK3 GluK4 GluK5 |
GRIK1 GRIK2 GRIK3 GRIK4 GRIK5 |
GLUK5, GluR5, GluR-5, EAA3 GLUK6, GluR6, GluR-6, EAA4 GLUK7, GluR7, GluR-7, EAA5 GLUK1, KA1, KA-1, EAA1 GLUK2, KA2, KA-2, EAA2 |
| NMDA | GluN |
GluN1 NRL1A NRL1B |
GRIN1 GRINL1A GRINL1B |
GLUN1, NMDA-R1, NR1, GluRξ1 |
|
GluN2A GluN2B GluN2C GluN2D |
GRIN2A GRIN2B GRIN2C GRIN2D |
GLUN2A, NMDA-R2A, NR2A, GluRε1 GLUN2B, NMDA-R2B, NR2B, hNR3, GluRε2 GLUN2C, NMDA-R2C, NR2C, GluRε3 GLUN2D, NMDA-R2D, NR2D, GluRε4 |
||
|
GluN3A GluN3B |
GRIN3A GRIN3B |
GLUN3A, NMDA-R3A, NMDAR-L, chi-1 GLU3B, NMDA-R3B |
||
| ‘Orphan’ | (GluD) |
GluD1 GluD2 |
GRID1 GRID2 |
GluRδ1 GluRδ2 |
AMPA受容体

α-アミノ-3-ヒドロキシ-5-メチル-4-イソオキサゾールプロピオン酸受容体 (AMPA受容体、またはキスカル酸型受容体としても知られている) は、中枢神経系 (CNS) での高速シナプス伝達を媒介するグルタミン酸の非NMDA型イオンチャネル型 膜貫通受容体である。その名前は、人工グルタミン酸アナログAMPAによって活性化される能力に由来している。この受容体は、天然に存在するアゴニストであるキスカル酸にちなんでワトキンスらによって「キスカル酸型受容体」と最初に命名され、その後、コペンハーゲンのデンマーク王立薬科大学のTage Honoreらによって開発された選択的アゴニストにちなんで「AMPA受容体」という標識が付与された。AMPARは脳の多くの部分に見られ、神経系で最もよく見られる受容体である。AMPA受容体GluA2 (GluR2) 四量体は、最初に結晶化されたグルタミン酸受容体イオンチャネルである。
リガンド:
- 作動薬 : グルタミン酸, AMPA , 5‐フルオロウィラルジイン , ドウモイ酸 , キスカル酸など
- 拮抗薬 : CNQX , キヌレン酸 , NBQX , ペランパネル , ピラセタムなど
- 陽性アロステリック修飾薬: アニラセタム , シクロチアジド , CX-516 , CX-614など
- 陰性アロステリック修飾薬 : エタノール , ペランパネル , タランパネル , GYKI-52,466など
NMDA受容体

N-メチル-D-アスパラギン酸受容体 (NMDA受容体) は、イオンチャネル型グルタミン酸受容体の一種であり、グルタミン酸とコアゴニスト (すなわちD-セリンまたはグリシンのいずれか) の同時結合によってゲート開閉されたリガンド依存性イオンチャネルである。研究では、NMDA受容体がシナプス可塑性と記憶の調節に関与していることが示されている。
「NMDA受容体」という名前は、リガンドN-メチル-D-アスパラギン酸 (NMDA) に由来しており、これらの受容体で選択的アゴニストとして作用する。NMDA受容体が2つのコアゴニストの結合によって活性化されると、カチオンチャネルが開き、細胞内にNa+とCa2+が流入し、細胞の膜電位が上昇する。このように、NMDA受容体は興奮性受容体である。静止膜電位では、Mg2+やZn2+が受容体の細胞外結合部位に結合することで、NMDA受容体チャネルを通過するイオン流が遮断される。しかし、ニューロンが脱分極したとき、例えば、共局在化したシナプス後AMPA受容体の強い活性化によって、Mg2+による電位依存性ブロックが部分的に緩和され、活性化されたNMDA受容体を介したイオンの流入が可能になる。結果として生じるCa2+の流入は、さまざまな細胞内シグナル伝達カスケードを誘発し、最終的にはさまざまなキナーゼやホスファターゼの活性化を通じて神経細胞の機能を変化させる。
リガンド:
- 主要な内因性 共作動薬 : グルタミン酸およびD-セリンまたはグリシン
- 他の作動薬 : アミノシクロプロパンカルボン酸 (英語版) , D-シクロセリン , L-アスパラギン酸 , キノリネートなど
- 部分作動薬 : N-メチル-D-アスパラギン酸 (NMDA) , NRX-1074 (英語版) , 3,5-ジブロモ-L-フェニルアラニンなど
- 拮抗薬 : ケタミン , PCP , デキストロプロポキシフェン , ケトベミドン , トラマドール , キヌレン酸 (内因性)など
GABA受容体
GABA受容体は、動物の大脳皮質の主要な介在神経細胞で発現する主要な抑制性神経伝達物質である。
GABAA受容体

GABAA受容体は、リガンド依存性イオンチャネルである。これらの受容体の内因性リガンドであるGABA (γ-アミノ酪酸) は、中枢神経系における主要な抑制性神経伝達物質である。活性化されると、神経細胞へのCl-の流れを仲介し、神経細胞を過分極化する。GABAA受容体は、神経系を持つすべての生物に存在する。それらは哺乳類の神経系内に広く分布しているため、実質的にすべての脳機能で役割を果たしている。
さまざまなリガンドがGABAA受容体に特異的に結合し、Cl-チャネルを活性化または阻害することができる。
リガンド:
5-HT3受容体
五量体 5-HT3受容体は、ナトリウム(Na)イオン、カリウム(K)イオン、およびカルシウム(Ca)イオンに対して透過性がある。
ATP依存性チャネル

ATP依存性チャネルは、ヌクレオチド ATPとの結合に応答して開く。これらのチャネルは、サブユニットごとに2つの膜貫通ヘリックスを持つ三量体を形成し、細胞内側にはC末端とN末端の両方がある。
| Type | Class | IUPHAR-recommended protein name |
Gene | Previous names |
|---|---|---|---|---|
| P2X | N/A |
P2X1 P2X2 P2X3 P2X4 P2X5 P2X6 P2X7 |
P2RX1 P2RX2 P2RX3 P2RX4 P2RX5 P2RX6 P2RX7 |
P2X1 P2X2 P2X3 P2X4 P2X5 P2X6 P2X7 |
PIP2依存性チャネル
ホスファチジルイノシトール4,5-ビスリン酸 (PIP2) は、内向き整流性カリウムチャネル (Kir) に結合し、直接活性化する。PIP2は細胞膜脂質であり、イオンチャネルのゲート開閉における役割は、この分子の新しい役割を表している。
間接的調節
リガンド依存性イオンチャネルとは対照的に、受容体とイオンチャネルが単一分子ではなく、細胞膜内の別個のタンパク質である受容体系も存在する。この場合、イオンチャネルは直接開閉されるのではなく、受容体の活性化によって間接的に調節される。
Gタンパク質結合型受容体

Gタンパク質結合型受容体は、別名でGタンパク質共役型受容体、7回膜貫通型受容体、7TM受容体とも呼ばれる受容体の大規模なタンパク質ファミリーを構成し、細胞外の分子を感知し、細胞内のシグナル伝達経路を活性化し、最終的には細胞応答を活性化する。それらは細胞膜を7回通過する。Gタンパク質結合型受容体は、数百ものメンバーが同定されている巨大なファミリーである。イオンチャネル結合型受容体 (GABAB、NMDAなど) はそれらの一部にすぎない。
表1. 三量体Gタンパク質の3つの主要なファミリー
| FAMILY | SOME FAMILY MEMBERS | ACTION MEDIATED BY | FUNCTIONS |
|---|---|---|---|
| I | GS | α | アデニリルシクラーゼを活性化し、Ca2+チャネルを活性化 |
| Golf | α | 嗅覚ニューロンのアデニル酸シクラーゼを活性化 | |
| II | Gi | α | アデニリルシクラーゼを阻害 |
| βγ | K+チャンネルを活性化 | ||
| G0 | βγ | K+チャネルを活性化。 Ca2+チャネルを不活性化 | |
| α and βγ | ホスホリパーゼC-βを活性化 | ||
| Gt (transducin) | α | 脊椎動物の桿体視細胞における環状GMPホスホジエステラーゼを活性化 | |
| III | Gq | α | ホスホリパーゼC-βを活性化 |
GABAB受容体
GABAB受容体は、γ-アミノ酪酸の代謝型膜貫通受容体である。それらは、Gタンパク質を介してK+チャネルと結合しており、活性化すると細胞内の電位を低下させ、過分極効果を発揮する。
リガンド:
- 作動薬 : GABA , バクロフェン , γ-ヒドロキシ酪酸 , フェニブトなど。
- 陽性アロステリック修飾薬 : CGP-7930 , フェンディリン 、 BSPPなど。
- 拮抗薬 : 2-OH-サクロフェン , サクロフェン , SCH-50911
Gαシグナル伝達
環状アデノシン一リン酸 (cAMP) 生成酵素アデニル酸シクラーゼは、Gαs経路とGαi/o経路の両方のエフェクターである。哺乳類の10種類のAC遺伝子産物は、組織分布および/または機能に微妙な違いがあり、すべて細胞基質のアデノシン三リン酸 (ATP) のcAMPへの変換を触媒し、すべてGαsクラスのGタンパク質によって直接刺激される。逆に、Gαi/o型のGαサブユニットとの相互作用は、ACによるcAMPの生成を阻害する。このように、Gαs に結合した GPCR は Gαi/o に結合した GPCR の作用を相殺し、逆もまた同様である。その後、細胞基質のcAMPのレベルにより、さまざまなイオンチャネルの活性だけでなく、セリン/スレオニン特異的プロテインキナーゼ A (PKA) ファミリーのメンバーが決定される。その結果、cAMPはセカンドメッセンジャーと見なされ、PKAはセカンダリエフェクターであると見なされる。
Gαq/11経路のエフェクターはホスホリパーゼC-β (PLCβ) であり、膜結合型ホスファチジルイノシトール4,5-ビスリン酸 (PIP2) のセカンドメッセンジャーであるイノシトール1,4,5三リン酸 (IP3) およびジアシルグリセロール (DAG) への開裂を触媒する。IP3は小胞体 (ER) の膜にあるIP3受容体に作用して小胞体からのCa2+放出を誘発し、DAGは原形質膜に沿って拡散し、プロテインキナーゼC (PKC) と呼ばれる第二のセリン/スレオニンキナーゼの細胞膜局在性を活性化する。PKCの多くのアイソフォームは細胞内Ca2+の増加によっても活性化されるため、これらの両方の経路がお互いに収束して、同じ二次エフェクターを介して信号を送ることもできる。細胞内Ca2+の上昇は、カルモジュリンと呼ばれるタンパク質にも結合し、アロステリックに活性化されるが、このタンパク質はCa2+/カルモジュリン依存性キナーゼ (CAMK) などの酵素と結合し、アロステリックに活性化する。
Gα12/13経路のエフェクターは3つのRhoGEF (p115-RhoGEF、PDZ-RhoGEF、LARG) であり、Gα12/13に結合すると、細胞質の低分子量GTPアーゼ、Rhoをアロステリックに活性化する。Rhoは一旦GTPに結合すると、その後、Rhoキナーゼ (ROCK) などの細胞骨格調節に関与するさまざまなタンパク質を活性化することができる。Gα12/13に結合するほとんどのGPCRは、他のサブクラス、多くの場合Gαq/11にも結合する。
Gβγシグナル伝達
上記の説明は、特に活性化されたGαi/o共役GPCRの場合には重要になる可能性があるGβγシグナル伝達の影響を無視している。Gβγの主なエフェクターは、Gタンパク質調節内向き整流性K+チャネル (GIRKs)、P/Q-およびN型電位依存性Ca2+チャネル、ならびにACおよびPLCのいくつかのアイソフォーム、およびいくつかのホスホイノシチド3キナーゼ (PI3K) アイソフォームなどの様々なイオンチャネルである。
臨床関連性
リガンド依存性イオンチャネルは、麻酔薬とエタノールが効果を発揮する主要な部位である可能性があるが、これについて明確な証拠はまだ確立されていない。特に、GABA受容体およびNMDA受容体は、臨床麻酔で使用される濃度と同様の濃度で麻酔薬の影響を受ける。
その機構を理解し、それらの受容体上で機能しうる化学的/生物学的/物理的な要素を探求することにより、予備実験やFDAによりますます多くの臨床応用が証明されている。
- メマンチンは、中等度から重度のアルツハイマー病の治療薬として、米国食品医薬品局および欧州医薬品庁から承認されており、現在、英国国立保健医療技術研究所 (National Institute for Health and Care Excellence) から、他の治療法が失敗に終わった患者のために限定的な推奨を受けている。
- 抗うつ薬治療
- アゴメラチンは、メラトニン作動性(melatonergic)-セロトニン作動性(serotonergic)の二重経路に作用する薬剤の一種であり、臨床試験中に不安うつ病の治療に有効性が示されている。研究ではまた、非定型うつ病およびメランコリー型うつ病の治療における有効性を示唆している。
関連項目
- 受容体 (生化学)
- 活動電位
- 電位依存性カルシウムチャネル
- カルシウム活性化カリウムチャネル
- 環状ヌクレオチド感受性イオンチャネル
- 酸感受性イオンチャネル
- リアノジン受容体
- イノシトール三リン酸受容体
参考文献
外部リンク
- European Bioinformatics Institute Ligand-Gated Ion Channelデータベース . 2007年4月11日確認済.
- “Revised Recommendations for Nomenclature of Ligand-Gated Ion Channels”. IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology. 2020年8月2日閲覧。
- www.esf.edu
- www.genenames.org
- “1.A.9 The Neurotransmitter Receptor, Cys loop, Ligand-gated Ion Channel (LIC) Family”. tcdb.org. Saier Lab Bioinformatics Group. 2020年8月3日閲覧。