
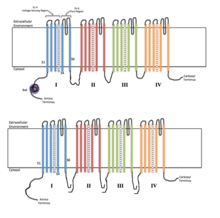
 4つの相同なドメインまたはサブユニットがイオンチャネルを構成する。S4電位センサーセグメントが+で示されている。
| |
| 識別子 | |
|---|---|
| 略号 | VIC |
| Pfam clan | CL0030 |
| TCDB | 1.A.1 |
| OPM superfamily | 8 |
| OPM protein | 2a79 |

電位依存性イオンチャネル(でんいいぞんせいイオンチャネル、英: voltage-gated ion channel)は、チャネル近傍の膜電位の変化によって活性化されるイオンチャネルである。膜電位はチャネルタンパク質のコンフォメーションを変化させ、チャネルの開閉を調節する。一般的に細胞膜はイオンを透過させないため、イオンは膜貫通タンパク質のチャネルを通って拡散する必要がある。電位依存性イオンチャネルは神経組織や筋組織など興奮性細胞で重要な役割を果たし、電位変化に応答した迅速かつ協調的な脱分極を可能にする。電位依存性イオンチャネルは軸索やシナプス上に存在し、方向性を持って電気信号を伝達する。電位依存性イオンチャネルには通常イオン特異性が存在し、ナトリウムイオン(Na+)、カリウムイオン(K+)、カルシウムイオン(Ca2+)、塩化物イオン(Cl-)にそれぞれ特異的なチャネルが同定されている。チャンネルの開閉は、細胞膜の両側のイオン濃度、すなわち電荷勾配の変化によって引き起こされる。
構造
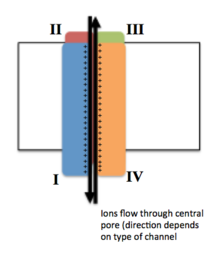
一般的に、電位依存性イオンチャネルは中心部にポアが形成されるように配置されたいくつかのサブユニットから構成され、イオンは電気化学的勾配に従ってポアを通って移動する。チャネルにはイオン特異性が存在する傾向があるが、似たサイズで似た電荷を持つ異なるイオンが通過することもある。電位依存性イオンチャネルの機能は、電位センサー、ポア、ゲートという3つの主要なユニットが担っている。Na+、K+、Ca2+チャネルは中心部にポアを形成するように配置された4つの膜貫通ドメインから構成される。大部分のNa+、Ca2+チャネルの場合はこれら4つのドメインは単一のαサブユニットの一部であるが、大部分のK+チャネルでは4つのαサブユニットが膜貫通ドメイン1つずつを構成している。膜貫通セグメントはS1からS6と表記され、これらはすべてαヘリックスを形成し、それぞれ専門的機能を有する。S5、S6とポアループはイオンの透過に主要な役割を果たし、チャネルのゲートとポアを構成する。一方、S1からS4は電位センサー領域を構成する。K+チャネルの4つのサブユニットは同一のものである場合も、異なるものである場合もある。中心となる4つのαサブユニットに加えて、酸化還元酵素活性を持ち、チャネル機能の調節を行うβサブユニットが存在する場合もある。βサブユニットは細胞膜の内側表面に位置し、膜を貫通していない。βサブユニットは小胞体でαサブユニットと共に組み立てられる。
機構
カリウムチャネルの結晶学的構造研究からは、膜に電位差が生じると、それに伴う電場によってカリウムチャネルのコンフォメーション変化が誘導される。このコンフォメーション変化によってチャネルが開き、膜を越えたイオンの流入や流出に十分なほどチャネルタンパク質の形状が変化する。この濃度勾配に従ったイオンの移動によって、細胞膜の脱分極に十分な電流が発生する。
電位依存性ナトリウムチャネルとカルシウムチャネルは、4つの相同なドメインを持つ1つのポリペプチドから構成される。各ドメインには6本の膜貫通αヘリックスが存在する。これらのヘリックスのうち、S4が電位センサーヘリックスである。S4セグメントには多くの正電荷が存在し、細胞外の高い正電荷と反発してチャネルの閉状態が維持される。
一般的に、イオンチャネルの電位センサー領域は膜を挟んだ電位の変化の検知を担い、チャネルの開閉を開始する。S1からS4のαヘリックスが一般的にこの役割を担っている。カリウムチャネルとナトリウムチャネルでは、電位センサーであるS4ヘリックスの反復モチーフには正に帯電したリジンやアルギニンが含まれている。静止状態では、各S4ヘリックスの半分は細胞質と接触している。脱分極が起こると、S4ドメインの正に帯電した残基は、膜の細胞外表面に向かって移動する。最初の4つのアルギニンがゲート電流を担っていると考えられており、膜の脱分極に応答したチャネルの活性化に伴って、細胞外の溶媒に向かって移動する。タンパク質に結合した10–12個の正電荷が移動することで、チャネルを開くための構造変化が引き起こされる。この移動が起こる正確な機構に関してこれまでのところ合意は得られていないが、現在の理論の例としては、カノニカル(canonical)モデル、トランスポーター(transporter)モデル、パドル(paddle)モデル、ツイスト(twisted)モデルなどが提唱されている。
これらのチャネルの電位センサードメインの主要な機能的領域には一般的にS3bとS4ヘリックスからなる領域が含まれており、その形状から「パドル」と呼ばれているが、この領域はさまざまな細胞や生物種の間で交換可能な、保存された配列であるようである。同様の電位センサーパドルは、さまざまな生物種の電位依存性ホスファターゼのファミリーでも発見されている。火山に生息する古細菌の生物種のパドル領域をラット脳のカリウムチャネルに遺伝子工学的に導入すると、パドル全体の完全な交換が行われている限り、完全に機能するイオンチャネルが得られる。このような「モジュール性」によって、特徴付けが十分になされていない、高価である、調製が困難といった制限を受けることなく、単純で安価なモデルシステムを用いてこの領域の機能や疾患における役割、挙動を制御する医薬品の研究を行うことが可能となっている。
電位依存性イオンチャネルは一般的には膜の脱分極によって活性化されるが、内向き整流性カリウムチャネルなど一部のチャネルは過分極によって活性化される。
ゲートはチャネルの電位センサー領域と共役していると考えられており、イオンの流れに対する機械的な障害物となる部分が含まれるようである。この障害物として機能するセグメントはS6ドメインであると考えられているが、その正確な機構は不明である。考えられる機構としては、S6セグメントがはさみのような動きをしてイオンを透過させる、S6セグメントが2つのセグメントに分かれてイオンを透過させる、あるいはS6自体がゲートの役割を果たしている、といったものがある。S4セグメントの動きがS6に影響を与える機構はまだ解明されていないが、S4-S5リンカーの動きによってS6が開くという仮説が立てられている。
イオンチャネルの不活性化は開口から数ミリ秒以内に起こる。不活性化は、ポアの開口を制御する細胞内ゲートによって媒介されると考えられている。このゲートは、ball-and-chainモデルとしてモデル化されている。不活性化の際には、チェーンが折りたたまれ、ボールがチャネルを通過するイオンの流れを遮断する。速い不活性化はS4セグメントの膜内移動による活性化と直接的に関係しているが、S4の移動と不活性化ゲートの作動をつなぐ機構は不明である。
タイプ
ナトリウムチャネル
ナトリウムチャネルは多くの異なる細胞種で類似した機能を持つ。ヒトではナトリウムチャネルをコードする遺伝子が10種類同定されているが、一般的にそれらの機能は異なる生物種や細胞種の間で保存されている。
カルシウムチャネル
ヒトではカルシウムチャネルの遺伝子は16種類同定されているが、このタイプのチャネルは細胞種によって機能が異なる。一部の神経細胞では、カルシウムチャネルはナトリウムチャネルと同様に活動電位を形成する。カルシウムチャネルはシナプス前神経終末において、神経伝達物質の放出にも関与している。大部分の細胞では、カルシウムチャネルは細胞内のカルシウム濃度の制御に関与しており、さまざまな生化学的過程を調節する。
カリウムチャネル
カリウムチャネルは電位依存性チャネルの中で最大かつ最も多様なクラスであり、ヒトではカリウムチャネルをコードする遺伝子は100以上存在する。これらのチャネルはゲート機能が大きく異なり、不活性化が極めて遅いものや迅速に不活性化するものなどがある。こうした活性化時間の差異は活動電位の発火の持続時間や速度に影響を与え、軸索に沿った電気伝導やシナプス伝達に大きな影響を持つ。カリウムチャネルは構造面で他のチャネルとは異なる。カリウムチャネルは4つの異なるポリペプチドサブユニットを含んでいるのに対し、他のチャネルは4つの相同なドメインを持つ単一のポリペプチドから構成される。
塩素チャネル
塩素チャネルは、すべての神経細胞種に存在する。塩素チャネルの主要な役割は神経細胞の興奮の制御であるが、細胞の静止電位の維持にも寄与し、細胞体積の調節も補助する。
プロトンチャネル
電位依存性プロトンチャネルは、ヒドロニウムの形態の水素イオンを介して電流を流し、pH依存的な脱分極によって活性化される。細胞内の酸を除去する機能を持つ。
系統学
細菌で発現しているタンパク質の系統学的研究により、電位依存性ナトリウムチャネルスーパーファミリーの存在が明らかにされている。その後の研究により、内向き整流性カリウムチャネル、イノシトール-1,4,5-トリスリン酸/リアノジン受容体型カルシウムチャネル(en:Ryanodine-Inositol 1,4,5-triphosphate receptor calcium channels)、TRPチャネル、ポリスシスチン型カチオンチャネル(en:Polycystin cation channel family)、イオンチャネル型グルタミン酸受容体、カルシウム依存性塩素チャネル、1価カチオン:プロトンアンチポーター1(en:Monovalent cation:proton antiporter-1)、カリウムトランスポーターなど、他のさまざまなイオンチャネルやトランスポーターが系統学的に電位依存性イオンチャネルと関係していることが示されている。