
| 配列および環境情報のみに基づいてポリペプチド配列の二次構造、三次構造、四次構造を予測することは可能か?逆タンパク質フォールディング問題: 特定の環境条件下で所与の構造を採用することになるポリペプチド配列を設計することは可能か? これは近年いくつかの小球状タンパク質で達成されている。 |
|
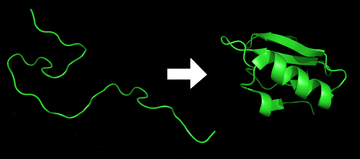

タンパク質フォールディング (英語: Protein folding) とは、タンパク質鎖がその本来の三次元構造、通常は生物学的に機能するコンホメーション(立体構造)を、迅速かつ再現性のある方法で獲得する物理的なプロセスである。これは、ポリペプチドがランダムコイルからその特徴的で機能的な三次元構造に折りたたまれる物理的な過程である。それぞれのタンパク質は、mRNAの配列からアミノ酸の直鎖に翻訳されるとき、折りたたまれていないポリペプチドまたはランダムコイルとして存在する。そのポリペプチドは、安定した (長続きする) 立体構造を欠いている (第1図の左側)。そのポリペプチド鎖がリボソームで合成されていく過程で、直鎖が三次元構造に折りたたまれる。フォールディングは、ポリペプチド鎖の翻訳中でも始まる。アミノ酸は互いに相互作用して、明確に定義された三次元構造、つまり天然状態として知られている折りたたまれたタンパク質 (図の右側) を生成する。結果として生じる三次元構造は、アミノ酸配列または一次構造 (アンフィンセンのドグマ) によって決定される。
タンパク質が機能を発揮するために正しい三次元構造が不可欠であるが、機能性タンパク質の一部は折りたたまれていない状態のままになっていることがあり、ゆえにタンパク質動力学が重要となる。本来の構造に折りたたまれないと、一般に不活性なタンパク質が生成されるが、場合によっては、誤って折りたたまれたタンパク質の機能が改変されたり、毒性のある機能性を持つこともある。いくつかの神経変性疾患やその他の疾患は、誤って折りたたまれたタンパク質によって形成されたアミロイド原線維の蓄積に起因すると考えられている。多くのアレルギーは、一部のタンパク質が正しく折りたたまれていないことが原因で、免疫系が特定のタンパク質構造に対する抗体を産生しないために引き起こされる。
タンパク質の変性は、折りたたまれた状態から折りたたまれていない状態に移行するプロセスである。これは、調理、火傷、プロテオパチー、その他の状況で起こる。
フォールディング・プロセスの所要時間は、目的のタンパク質によって劇的に異なる。細胞外で調べたとき、最も遅く折りたたまれるタンパク質は、主にプロリン異性化のために折りたたまれるのに数分から数時間を要し、プロセスが完了するまでにチェックポイントのようないくつかの中間状態を通過する必要がある。一方、長さが100アミノ酸までの非常に小さなシングルドメインタンパク質は、通常、1回のステップで折りたたむことができる。時間スケールはミリ秒が一般的で、非常に速い既知のタンパク質のフォールディング反応は数マイクロ秒以内に完了する。
タンパク質のフォールディング過程
一次構造
タンパク質の一次構造である直線的なアミノ酸配列は、その本来のコンホメーションを決定する。特定のアミノ酸残基と、ポリペプチド鎖内におけるそれらの位置は、タンパク質のどの部分が密接に折り重なり、その三次元構造を形成するかを決定する要因となる。アミノ酸組成は配列ほど重要ではない。しかし、フォールディングの本質的な事実は、各タンパク質のアミノ酸配列が、本来の構造とその状態に到達するための経路の両方を指定する情報を含んでいることである。これは、ほぼ同じアミノ酸配列が常に同じように折りたためるということではない。類似したタンパク質でも、環境要因によってコンホメーションは異なり、どこで見つかったかによって異なる方法で折りたたまれる。
二次構造
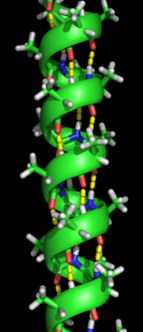

二次構造の形成は、タンパク質がその本来の構造を取るための、フォールディング・プロセスの最初のステップである。二次構造の特徴はαヘリックスやβシートとして知られている構造で、ライナス・ポーリングによって最初に述べられたように、分子内水素結合によって安定化されているために急速に折りたたまれる。分子内水素結合の形成は、タンパク質の安定性に別の重要な貢献をしている。αヘリックスは、主鎖の水素結合により螺旋状に形成される (右図参照)。βプリーツシートは、水素結合を形成するために主鎖が折り曲がって形成される構造である (左図参照)。水素結合は、ペプチド結合のアミド水素とカルボニル酸素の間にある。逆平行βプリーツシートと平行βプリーツシートが存在し、平行βシートによって形成される傾斜水素結合と比較して、逆平行βプリーツシートの方が理想的な180度の角度で水素結合を形成するため水素結合の安定性が高くなっている。
三次構造
αヘリックスおよびβプリーツシートは、本質的に両親媒性であるか、または親水性部分と疎水性部分を含むことができる。二次構造のこの性質はタンパク質の三次構造形成を助け、親水性側がタンパク質を取り囲む水溶性環境に面し、疎水性側がタンパク質の疎水性コアに面するようにフォールディングが起こる。二次構造は階層的に三次構造の形成によって取って代わられる。タンパク質の三次構造が形成され、疎水性相互作用によって安定化されると、2つのシステイン残基間に形成されたジスルフィド結合の形で共有結合が存在することもある。タンパク質の三次構造には、単一のポリペプチド鎖が含まれるが、折りたたまれたポリペプチド鎖の追加の相互作用により四次構造が形成される。
四次構造
三次構造は、いくつかのタンパク質の四次構造の形成で取って代わられ、これには通常、すでに折りたたまれてたサブユニットの「集合」または「会合」を含む。言い換えれば、複数のポリペプチド鎖が相互作用して、完全に機能する四次タンパク質を形成する可能性がある。
タンパク質フォールディングの推進力
フォールディングは、主に疎水性相互作用、分子内水素結合の形成、ファンデルワールス力によって導かれる自発的過程であり、配座エントロピーによって対抗を受ける。フォールディングのプロセスは、多くの場合、共翻訳的(部分的な翻訳)に開始されるため、タンパク質のC末端部分がまだリボソームによって合成されている間に、先に翻訳されたタンパク質のN末端が折りたたみ始まる。ただし、タンパク質分子は、生合成中または生合成後に自発的に折りたたまれることがある。これらの高分子は「自分自身で折りたためる」と考えられるが、このプロセスは溶媒 (水または脂質二重層)、塩の濃度、pH、温度、補助因子および分子シャペロンの存在にも依存している。
タンパク質は、可能な曲げ角度またはコンホメーションが制限されているため、フォールディング能力に制限がある。タンパク質フォールディングのこれらの許容角度は、ラマチャンドランプロットとして知られている二次元プロットで表され、許容回転のpsiとphi角度で示される。
疎水性効果
タンパク質のフォールディングが自発的な反応であるためには、細胞内で熱力学的に有利でなければならない。タンパク質のフォールディングは自発的な反応であることが知られているため、負のギブス自由エネルギー値をとる必要がある。タンパク質のフォールディングにおけるギブス自由エネルギーはエンタルピーとエントロピーに直接関係している。負のデルタGが発生し、タンパク質のフォールディングが熱力学的に有利になるためには、エンタルピー、エントロピー、またはその両方が有利でなければならない。
水にさらされる疎水性側鎖の数を最小限にすることは、フォールディングプロセスの背後にある重要な推進力である。疎水効果とは、タンパク質の疎水性鎖がタンパク質の中心部 (親水性環境から離れる) に凝集する現象である。水性環境下では、水分子はタンパク質の疎水性領域や側鎖の周囲に凝集し、秩序だった水分子の水殻を形成する傾向がある。疎水性領域を中心とした水分子の秩序は系内の秩序を増大させ、エントロピーの負の変化に寄与する (系内のエントロピーは減少する)。水分子はこれらの水分子ケージに固定されており、疎水性凝集、または疎水性基の内側へのフォールディングを促進する。疎水性凝集は、秩序だった水分子を解放する水ケージの破壊を介してシステムにエントロピーを導入する。球状に折りたたまれたタンパク質のコア内で相互作用する多数の疎水性基は、膨大に蓄積されたファンデルワールス力 (特にロンドン分散力) のため、折りたたみ後のタンパク質の安定性に大きく貢献する。疎水性効果は、大きな疎水性領域を含む両親媒性分子を含む水性媒体が存在する場合にのみ、熱力学の推進力として存在する。水素結合の強さは環境に依存するため、疎水性コアに包まれた水素結合は、水性環境にさらされた水素結合よりも天然状態の安定性に寄与する。
球状の折りたたみを持つタンパク質では、疎水性アミノ酸はランダムに分布したり、一緒にクラスター化されるのではなく、一次配列に沿って散在する傾向がある。しかし、天然変成傾向のある新生(de novo)タンパク質は、一次配列に沿って疎水性アミノ酸がクラスター化するという逆のパターンを示す。
シャペロン

分子シャペロン (英: molecular chaperone) は、生体内 (in vivo) で他のタンパク質を正しく折りたたむのに役立つタンパク質の一種である。シャペロンは、すべての細胞内区画に存在し、ポリペプチド鎖と相互作用して、タンパク質の本来の三次元コンホメーションを形成できるようにする。ただし、シャペロン自体は、それらが補助しているタンパク質の最終構造には含まれていない。シャペロンは、新生ポリペプチドがリボソームによって合成されている場合でも、フォールディングを助けることができる。分子シャペロンは、結合することによって機能し、フォールディング経路でタンパク質の不安定な構造を安定化させるが、シャペロンには、それらが補助しているタンパク質の正しい本来の構造を知るために必要な情報は含まれておらず、むしろ、シャペロンは、誤った折りたたみ構造を防ぐことによって機能する。このように、シャペロンは実際には、本来の構造に向かうフォールディング経路に関与する個々のステップの速度を増加させることはなく、その代わりに、適切な中間体の探索を遅くする可能性のあるポリペプチド鎖の不要な凝集を減らすことで機能し、ポリペプチド鎖が正しいコンホメーションをとるためのより効率的な経路を提供する。シャペロンは、フォールディング触媒と混同されるべきではなく、フォールディング経路の遅いステップを実際に触媒する。フォールディング触媒の例は、タンパク質ジスルフィド異性化酵素(protein disulfide isomerases)およびペプチジルプロリル異性化酵素(peptidyl-prolyl isomerases)があり、それぞれジスルフィド結合の形成またはシスおよびトランス立体異性体間の相互変換に関与している可能性がある。シャペロンは、生体内でのタンパク質フォールディングのプロセスにおいて重要であることが示されている。なぜなら、シャペロンは、タンパク質が「生物学的に適切な」状態になるために十分効率的に適切な配列とコンホメーションをとるのに必要な手助けタンパク質に提供するからである。これは、in vitroで行われたタンパク質フォールディング実験で実証されたように、ポリペプチド鎖は理論的にはシャペロンの助けなしにその本来の構造に折りたたむことができることを意味するものであるが、このプロセスはあまりにも非効率的であるか、または遅すぎて生物学的システムには存在しないことが判明している。したがって、シャペロンは生体内でのタンパク質のフォールディングに必要である。シャペロンは、本来構造の形成を助ける役割に加えて、タンパク質の輸送、分解、さらには外部変性因子にさらされた変性タンパク質が正しい本来の構造にリフォールディング(再折りたたみ)する機会を与えるなど、様々な役割に関与していることが明らかになっている。
完全に変性したタンパク質は、三次構造と二次構造の両方を欠いており、いわゆるランダムコイルとして存在している。特定の条件下では、一部のタンパク質は再折りたたみ(リフォールド)する可能性があるが、多くの場合、変性は不可逆的である。細胞は、熱変性の影響からタンパク質を守るために、熱ショックタンパク質 (シャペロンの一種) として知られる酵素を用いて、他のタンパク質のフォールディングや折りたたまれた状態の維持を助けている。熱ショックタンパク質は、細菌からヒトにいたるまで、調査したすべての種で発見されており、非常に早い段階で進化し、重要な機能を持っていることが示唆されている。一部のタンパク質は、シャペロンの助けを借りて他のタンパク質との相互作用によりフォールディングが中断されないように個々のタンパク質を分離するか、誤って折りたたまれたタンパク質を展開して本来の正しい構造にリフォールディングをしない限り、細胞内では全く折りたたまれないことがある。この機能は、不溶性のアモルファス凝集体への沈殿のリスクを防ぐために非常に重要である。タンパク質の変性や天然状態の破壊に関与する外部要因には、温度、外部磁場 (電場、磁場)、分子クラウディング、さらにはタンパク質のフォールディングに大きな影響を与える可能性のある空間の制限 (すなわち閉じ込め) が含まれる。高濃度の溶質、極端なpH、機械的な力、化学的変性剤の存在も同様にタンパク質の変性に寄与する。これらの個々の要因はストレスとしてまとめて分類される。シャペロンは、細胞ストレス時には高濃度で存在し、変性タンパク質や誤って折りたたまれたタンパク質だけでなく、新生タンパク質の適切なフォールディングを助けることが示されている。
一部の条件下では、タンパク質は生化学的に機能する形に折りたたまれない。細胞が通常生存する温度範囲よりも高い温度や低い温度では、熱的に不安定なタンパク質は折りたたまれなかったり、変性する (これが煮沸すると卵白が不透明になる理由である)。しかし、タンパク質の熱安定性が一定であるとは限らない。例えば、超好熱性細菌は122 °Cの高温で生育することが確認されているが、これにはもちろん、重要なタンパク質やタンパク質集合体の完全な補体が、その温度以上で安定している必要がある。
大腸菌はバクテリオファージT4の宿主であり、ファージにコードされたgp31タンパク質は、大腸菌のシャペロンタンパク質 GroESと機能的に相同であるように見え、感染時にバクテリオファージT4ウイルス粒子の組み立てにおいてそれを置き換えることができる。GroESと同様に、gp31はGroELシャペロニンとの安定な複合体を形成する。これはバクテリオファージT4メジャー・キャプシド・タンパク質gp23のin vivoでのフォールディングおよび組み立てに絶対的に必要である。
タンパク質の誤ったフォールディングと神経変性疾患
タンパク質は、通常の天然状態を得られない場合、ミスフォールド(誤った折りたたみ)をしていると考えられる。これは、アミノ酸配列の突然変異または外部要因による通常のフォールディングプロセスの混乱が原因である可能性がある。ミスフォールドされたタンパク質は、通常、クロスβ構造として知られる超分子配列で組織化されたβシートを含んでいる。これらのβシートが豊富に含んだ集合体は、非常に安定し極めて不溶性であり、一般にタンパク質分解に対して耐性がある。これらのフィブリル状集合体の構造的安定性は、βストランド間の主鎖水素結合によって形成されたタンパク質モノマー間の広範な相互作用によってもたらされる。タンパク質のミスフォールディングは、他のタンパク質の凝集体またはオリゴマーへのさらなるミスフォールディングや蓄積を引き起こす可能性がある。細胞内で凝集したタンパク質のレベルが上昇すると、変性疾患や細胞死を引き起こす可能性のあるアミロイド様構造の形成につながる。アミロイドは、分子間水素結合を含む線維状構造であり、極めて不溶性で、転換されたタンパク質の集合体から作られる。そのため、プロテアソーム経路では、凝集する前にミスフォールドしたタンパク質を分解するのに十分な効率が得られない場合がある。ミスフォールドされたタンパク質は、互いに相互作用して構造化された凝集体を形成し、分子間相互作用を通じて毒性を獲得する可能性がある。
凝集タンパク質は、クロイツフェルト・ヤコブ病、牛海綿状脳症 (狂牛病) などのプリオン関連疾患、アルツハイマー病および家族性アミロイド心筋症または多神経症などのアミロイド関連疾患、ならびにハンチントン病およびパーキンソン病などの細胞内凝集性疾患と関連している。これらの加齢性変性疾患は、不溶性の細胞外凝集体および(または)クロスβアミロイド原線維を含む細胞内封入体へのミスフォールドタンパク質の凝集に関連している。これは凝集体が原因なのか、それとも単にタンパク質の恒常性の喪失、合成、フォールディング、凝集、タンパク質代謝回転のバランスを反映しているだけなのかは完全には明らかではない。最近、欧州医薬品庁は、トランスサイレチンアミロイド疾患の治療のためのタファミディス(Tafamidis)またはビンダケル (Vyndaqel; 四量体トランスサイレチンの動態安定化剤) の使用を承認した。このことは、ヒトのアミロイド疾患において、アミロイド原線維形成プロセス (原線維自体ではなく) が、有糸分裂後の組織の変性を引き起こすことを示唆している。フォールディングや機能ではなく、ミスフォールディングや過度の分解は、アンチトリプシン関連肺気腫、嚢胞性線維症、リソソーム蓄積症などの多くのプロテオパチー疾患を引き起こし、機能の喪失が障害の根源となっている。後者の疾患を修正するためにタンパク質補充療法が歴史的に使用されてきたが、新たなアプローチは、薬理シャペロンを使用して変異タンパク質を折りたたんで機能させる状態にすることが挙げられる。
タンパク質のフォールディングを研究するための実験技術
タンパク質のフォールディングに関する推論は、突然変異研究 (英語版) を通じて行うことができるが、タンパク質のフォールディングを研究するための実験技術は、通常、タンパク質の段階的アンフォールディングやフォールディングと、標準的な非結晶学的技術を用いたコンホメーション変化の観察に依存している。
X線結晶構造解析

X線結晶構造解析は、折りたたまれたタンパク質の三次元構造を解読するための、効率的で重要な方法の一つである。X線結晶構造解析を行うためには、対象となるタンパク質が結晶格子内に配置されている必要がある。タンパク質を結晶格子内に配置するためには、結晶化に適した溶媒を用意し、溶液中で過飽和状態の純粋なタンパク質を得て、溶液中で結晶を析出させる必要がある。タンパク質が結晶化されると、X線ビームは結晶格子を介して集中することができ、ビームを回折したり、様々な方向にビームを外側に向けて発射したりする。これらの出射ビームは、内包されたタンパク質の特定の三次元構成に相関している。X線は、タンパク質の結晶格子内の個々の原子を取り囲む電子雲と特異的に相互作用し、識別可能な回折パターンを生成する。電子密度雲をX線の振幅を関連付けることによってのみ、このパターンを読み取ることができ、この方法を複雑にする位相や位相角の仮定を導くことになる。フーリエ変換という数学的基礎によって確立された関係がなければ、「位相問題」は回折パターン予測を非常に困難にする。多重同型置換のような新しい方法では、重金属イオンの存在を利用してX線をより予測可能な方法で回折させ、関与する変数の数を減らして位相の問題を解決している。
蛍光分光法
蛍光分光法は、タンパク質の折りたたみ状態を調べるための高感度な手法である。フェニルアラニン(Phe)、チロシン(Tyr)、トリプトファン(Trp)の3つのアミノ酸は固有の蛍光特性を持つが、TyrとTrpのみが量子収率が高く、良好な蛍光シグナルが得られるため、実験的に使用されている。TrpとTyrは共に280 nmの波長で励起されるのに対し、Trpだけは295 nmの波長で励起される。それらの芳香族性のため、TrpとTyr残基は、タンパク質の疎水性コア、2つのタンパク質ドメイン間の界面、またはオリゴマータンパク質のサブユニット間の界面に、完全または部分的に埋もれていることがよくある。この無極性環境では、これらの残基は高い量子収率を持ち、それゆえ高い蛍光強度を示す。タンパク質の3次構造や4次構造が破壊されると、これらの側鎖は溶媒の親水性環境にさらされるようになり、量子収率が低下して低い蛍光強度になる。Trp残基については、その最大蛍光発光の波長も環境に依存する。
蛍光分光法は、変性剤の値の関数として、蛍光発光強度または最大発光波長の変動を測定することにより、タンパク質の平衡アンフォールディングを特徴付けるために使用される。ここで変性剤は、化学分子 (尿素、塩酸グアニジニウム)、温度、pH、圧力などである。互いに異なるが離散的なタンパク質の状態、すなわち天然状態、中間状態、アンフォールド状態の間の平衡は、変性剤の値に依存するため、それらの平衡混合物の全体的な蛍光シグナルもこの値に依存する。このようにして、全体的なタンパク質シグナルを変性剤の値に関連付けるプロファイルが得られる。平衡アンフォールディングのプロファイルは、アンフォールディングの中間体を検出し、識別することを可能にする。Hugues Bedouelleによって一般的な方程式が開発され、そのようなプロファイルから、ホモマーまたはヘテロマーのタンパク質のアンフォールディング平衡を特徴づける熱力学的パラメータを三量体まで、および潜在的には四量体まで得ることができた。蛍光分光法は、ストップフローのような高速混合装置と組み合わせて、タンパク質のフォールディング動態を測定し、シェブロンプロットを生成し、Phi値分析 (英語版) を導出することができる。
円偏光二色性
円偏光二色性は、タンパク質のフォールディングを研究するための最も一般的で基本的なツールの一つである。円二色性分光法は、円偏光の吸収を測定する。タンパク質では、αヘリックスやβシートなどの構造は不斉であるため、このような光を吸収する。この光の吸収は、タンパク質アンサンブルの折りたたみ度合いのマーカーとして機能する。この技術は、変性剤濃度または温度関数としてこの吸収の変化を測定することにより、タンパク質の平衡アンフォールディングを測定するために使用されてきた。変性溶解は、タンパク質のm値、または変性剤依存性と同様にアンフォールディングの自由エネルギーを測定する。温度溶解は、タンパク質の変性温度(Tm)を測定する。蛍光分光法に関しては、円二色性分光法をストップフローなどの高速混合装置と組み合わせて、タンパク質のフォールディング動態を測定し、シェブロンプロットを生成することができる。
タンパク質の振動円二色性
最近開発されたタンパク質の振動円二色性 (vibrational circular dichroism; VCD) 技術は現在、フーリエ変換(FT)機器を用いており、非常に大きなタンパク質分子でも、溶液中のタンパク質構造を決定する強力な手段を提供する。このようなタンパク質のVCD研究は、タンパク質結晶のX線回折、重水(D2O)中のタンパク質溶液のFT-IRデータ、または第一原理量子計算と組み合わせて、円偏光二色性(CD)からは得られない明確な構造決定を提供することがよくある。
タンパク質核磁気共鳴分光法
タンパク質のフォールディングは、NMR分光法を使用して日常的に研究されており、例えば、その天然状態でのタンパク質の主鎖アミドプロトンの水素-重水素交換 (英語版) を監視することで、タンパク質の残基固有の安定性と全体的な安定性の両方を提供する。
二重偏光干渉法
二重偏光干渉法は、分子層の光学特性を測定する表面ベースの技術である。タンパク質のフォールディングを特徴づけるために使用される場合、タンパク質の単層の全体的なサイズとその密度をサブ・オングストーム分解能でリアルタイムに測定することによってコンホメーションを測定するが、タンパク質フォールディングの速度論のリアルタイム測定は、~10 Hzよりも遅いプロセスに限られている。円偏光二色性と同様に、フォールディングのための刺激は変性剤または温度である可能性がある。
高時間分解能でのフォールディングの研究
タンパク質のフォールディングの研究は、高速で時間分解技術の開発によって近年、大きく進展した。実験者は、折りたたまれていないタンパク質のサンプルのフォールディングを迅速に誘発し、結果として生じるタンパク質動力学を観察する。使用されている高速化技術には、中性子散乱、超高速溶液混合、光化学的手法、レーザー温度ジャンプ分光法などがある。これらの技術の開発に貢献した多くの科学者の中には、Jeremy Cook, Heinrich Roder, Harry Gray, Martin Gruebele, Brian Dyer, William Eaton, Sheena Radford, Chris Dobson, Alan Fersht, Bengt Nölting, Lars Konermannがいる。
タンパク質分解
タンパク質分解 (proteolysis) は、広範囲の溶液条件下 (例: 高速並列タンパク質分解) でアンフォールドされた画分を探索するために日常的に使用されている。
単一分子力分光法
単離されたタンパク質やシャペロンを持つタンパク質のフォールディング機構を理解するために、光ピンセットやAFMなどの単一分子力技術 (Single-molecule force spectroscopy) が用いられてきた。光ピンセットは、単一のタンパク質分子をC末端とN末端から引き伸ばし、それを展開して、その後のリフォールディングを研究するために使用されてきた。この手法により、単一分子レベルでフォールディング率を測定できる。例えば、光ピンセットは最近、血液凝固に関与するタンパク質のフォールディングとアンフォールディングの研究に応用されている。ヴォン・ヴィレブランド因子 (von Willebrand factor (vWF)) は、血液凝固プロセスに不可欠な役割を持つタンパク質である。単一分子の光ピンセット測定を使用して、カルシウム結合vWFが血液中でせん断力センサーとして働くことを発見した。せん断力はvWFのA2ドメインのアンフォールディングにつながり、そのリフォールディング速度はカルシウムの存在下で劇的に向上する。最近では、単純な src SH3ドメインが、力を受けると複数のアンフォールディング経路にアクセスすることも明らかにされた。
ビオチン標識
ビオチン標識 (Biotin painting) は、(未)フォールディング・タンパク質の状態特異的な細胞スナップショットを可能にする。 ビオチン標識は、予測される天然変成タンパク質への偏りを示している。
タンパク質フォールディングの計算科学的研究
タンパク質フォールディングの計算科学的研究には、タンパク質の安定性、速度論、および構造予測に関連する3つの主要な側面が含まれる。以下の最近のレビューは、タンパク質フォールディングに利用可能な計算手法をまとめたものである。
レヴィンタールのパラドックス
1969年、サイラス・レヴィンタールは、折りたたまれていないポリペプチド鎖の自由度が非常に大きいため、タンパク質分子は天文学的な数の起こりうるコンホメーションを持っていることに着目した。彼の論文の中で、3300または10143という推定がなされている。レヴィンタールのパラドックスは、タンパク質が起こりうるすべてのコンホメーションを順次サンプリングして折りたたまれた場合、たとえコンホメーションが高速 (ナノ秒またはピコ秒スケール) でサンプリングされたとしても、天文学的な量の時間がかかるという観察に基づく思考実験である。タンパク質はこれよりもはるかに速く折りたたまれるという観測に基づいて、レヴィンタールは、ランダムなコンホメーション探索は発生しないため、タンパク質は一連の準安定な中間状態を経て折りたたまれなければならないと提案した。
タンパク質フォールディングのエネルギー地形
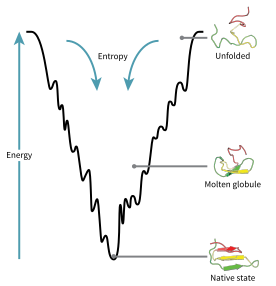
フォールディング中のタンパク質の配位空間は、エネルギー地形 (energy landscape) として可視化できる。Joseph BryngelsonとPeter Wolynesによると、タンパク質は最小フラストレーション原理に従っており、自然に進化したタンパク質はフォールディング時のエネルギー地形を最適化し、自然はタンパク質の折りたたみ状態が十分に安定するようにアミノ酸配列を選択していることを意味している。さらに、折りたたまれた状態の獲得は、十分に高速なプロセスにならなければならない。自然がタンパク質のフラストレーションのレベルを減らしたとしても、タンパク質のエネルギー地形における局所的な最小値が存在することからもわかるように、ある程度のフラストレーションは今のところ残っている。
これらの進化的に選択された配列の結果として、タンパク質は、天然状態に向かうグローバルな「ファンネル(漏斗)状のエネルギー地形」(José Onuchicの造語) を持っていると一般的に考えられている。この「フォールディング・ファンネル」地形により、タンパク質は、単一のメカニズムに限定されるのではなく、多数の経路や中間体のいずれかを介して天然状態にフォールディングできる。この理論は、モデルタンパク質の計算シミュレーション(格子タンパク質)と実験的研究の両方で支持されており、タンパク質構造の予測とタンパク質構造設計のための方法を改善するために使用されてきた。平準化自由エネルギー地形によるタンパク質フォールディングの説明も、熱力学第2法則と合致している。物理的には、エネルギー地形を、地理的な地形のように、単に最大値、鞍点、最小値、ファンネルを持った可視化可能なポテンシャル曲面や全エネルギー曲面の観点から考えることは、あるいはいくらか誤解を招く可能性がある。妥当な記述は、実際には、多様体が様々なより複雑な位相形態をとる可能性のある高次元の位相空間である。
折りたたまれていないポリペプチド鎖は、ファンネルの一番上に位置し、折りたたまれていないバリエーションの数が最も多く、エネルギー状態は高も高くなる。このようなエネルギー地形は、初期の可能性が多数あることを示しているが、可能なのは単一の天然状態のみである。しかし、それは可能な多くのフォールディング経路を明らかにしていない。同じ正確なタンパク質の異なる分子は、同じ天然構造に到達する限り、わずかに異なるフォールディング経路をたどり、異なる低エネルギー中間体を探すことができる場合がある。異なる経路は、各経路の熱力学的な有利性に応じて、異なる利用頻度を持つ可能性がある。これは、ある経路が他の経路よりも熱力学的に有利であることが分かった場合、本来の構造を追求するために、より頻繁に使用される可能性が高いことを意味する。タンパク質が折りたたみ始め、さまざまなコンホメーションをとると、常に以前よりも熱力学的に有利な構造を求め、エネルギーファンネルを通過し続けることになる。二次構造の形成は、タンパク質内の安定性の向上を強く示しており、ポリペプチド骨格によって想定される二次構造の一つの組み合わせだけが最低のエネルギーを持ち、ゆえにタンパク質は天然状態で存在することになる。ポリペプチドが折りたたみを開始すると形成される最初の構造の中には、αヘリックスおよびβターンがあり、αヘリックスはわずか100ナノ秒で形成され、βターンは1マイクロ秒で形成される。
エネルギー・ファンネル地形には、特定のタンパク質の遷移状態が見られる鞍点が存在する。エネルギー・ファンネル図の遷移状態とは、タンパク質が最終的に本来の構造をとることを想定した場合、そのタンパク質のすべての分子がとらなければならないコンホメーションである。どのタンパク質も、最初に遷移状態を通過しなければ、本来の構造をとることはできない。遷移状態は、単なる別の中間段階ではなく、天然状態の変化形または未熟な形と呼ぶことができる。遷移状態のフォールディングは律速であることが示されており、それが本来のフォールディングよりも高いエネルギー状態で存在しているとしても、本来の構造と極めて類似する。遷移状態の中には、タンパク質が折りたたむことができる核(足場)となる構造が存在しており、核の上に構造が段階的に完成してゆく「凝縮核形成」と呼ばれるプロセスによって形成される。
タンパク質フォールディングのモデリング

計算によるタンパク質構造予測のためのデ・ノボ (de novo) または第一原理 (ab initio) 的手法は、どちらもタンパク質フォールディングの実験的研究に関連しているが、厳密には区別されるものである。分子動力学法 (MD) は、タンパク質のフォールディングと動力学をイン・シリコ (in silico) で研究するための重要なツールである。最初の平衡フォールディング・シミュレーションは、暗黙の溶媒モデルとアンブレラ・サンプリング法を用いて行われた。計算コストが高いため、明示的な水を用いた第一原理計算によるフォールディング・シミュレーションは、ペプチドや非常に小さなタンパク質に限定される。より大きなタンパク質のMDシミュレーションは、実験的な構造の動力学、または、その高温アンフォールディングに限定される。小さなサイズのタンパク質 (約50残基以上) のフォールディングのような長時間のフォールディングプロセス (約1ミリ秒を超える) は、粗視化モデルを用いて解析することができる。
スタンフォード大学教授のビジェイ・S・パンデのグループが作成した100ペタFLOP級の分散コンピューティングプロジェクト Folding@home は、ボランティアのパーソナルコンピュータのCPUとGPUのアイドル処理時間を利用して、タンパク質のフォールディングをシミュレーションする。このプロジェクトは、タンパク質のフォールディングのミスフォールディング(誤った折りたたみ)を理解し、疾患研究のための創薬ドラッグデザインを加速することを目的としている。
D. E. Shaw Research社 (英語版) のカスタムASICと相互接続を中心に設計・構築された超並列スーパーコンピュータAnton(アントン)で、長時間の連続軌道シミュレーションが実行された。Antonを使用して実行されたシミュレーションの公開された最長の結果は、355KでのNTL9の2.936ミリ秒のシミュレーションである。
参考文献
- 藤博幸 編「タンパク質の立体構造入門」 講談社 (2010)
- Voet & Voet "Biochemistry" 3rd edition, Wiley (2004)
関連項目
- アントン (スーパーコンピュータ)
- シャペロン
- シェブロン・プロット
- 変性中間点
- ダウンヒル・フォールディング
- 高速並列タンパク質分解
- フォールディング (化学)
- Folding@Home
- Foldit - コンピュータ・ゲーム
- 力場 (化学)
- Pt-barrel
- タンパク質動力学
- タンパク質ミスフォールディング周期的増幅 (PMCA法)
- タンパク質構造予測ソフトウェア
- プロテオパチー
- Rosetta@home
- 分子力学モデリング用ソフトの比較
- 統計的ポテンシャル
- 時間分解質量分析法