
人類の進化(じんるいのしんか、英: human evolution)とは、他と異なる独立した生物種としての現生人類(英名:anatomically modern humans〈意:解剖学的現代人〉、学名:Homo sapiens〈ホモ・サピエンス〉、他説では Homo sapiens sapiens )を主題としながら、これが属する系統群(クレード)全体を見渡そうとする概念における、当該系統群が辿ってきた生物学的進化をいう。起源を紐解くという観点から、人類の起源(じんるいのきげん、英: human origins)などといった表現もある。
本項では、霊長類(サル目;霊長目)の出現を起点とし、人類の祖先系統を含む全ての化石人類、そして現生人類が出現してやがて唯一の残存種となるまでの、進化と絶滅の経緯についての解説を主題としながら、関連事象や研究史などについても言及する。
概要
人類の祖先に、どのような進化的変化が起きたかは、幅広い科学的探求の主題である。この研究は多くの分野、特に形質人類学、言語学、遺伝学、考古学などと関連している。
なお、「人類」という用語は人類の進化の文脈ではヒト科ヒト亜科ヒト族ヒト亜族ヒト属生物に対して用いられるが、他の属(アウストラロピテクス属など)を含むヒト亜族生物を指す場合もある。本記事では、人類という用語をチンパンジー亜族と分岐し直立二足歩行していたヒト亜族生物に用い、脳の発達したヒト属生物については学名で表記し、特にヒト属生物のうちホモ・サピエンス・サピエンスについては現生人類と表記する。
ヒト属(ホモ属)はおよそ200万年前にアフリカでアウストラロピテクス属から別属として分化し、ヒトの属するホモ・サピエンスは40万から25万年前に現れた。またこれらの他にも、すでに絶滅したヒト属の種が幾つか確認されている。その中にはアジアに生息したホモ・エレクトゥスや、ヨーロッパに生息したホモ・ネアンデルターレンシスが含まれる。
ホモ・サピエンスの進化と拡散については、アフリカ単一起源説と多地域進化説とが対立している(#人類進化のモデル)。アフリカ単一起源説では、アフリカで「最も近いアフリカの共通祖先 (RAO)」であるホモ・サピエンスが進化し、世界中に拡散してホモ・エレクトゥスとホモ・ネアンデルターレンシスに置き換わったとしている。多地域進化説を支持している科学者は世界中に分散した単一のヒト属、おそらくホモ・エレクトゥスが各地でそれぞれホモ・サピエンスに進化したと考えている。
化石の証拠はこの分野における激しい議論を解決するのに十分ではない。人類はホモ・ハビリスの頃から石器を使い始め、次第に洗練させてきた。およそ5万年前を境に現生人類の技術と文化は変わり始め、現代的行動がとられるようになった。
古人類学の歴史
古人類学は化石、道具のような遺物、居住の痕跡などにもとづく古代の人類研究である。現代的な科学としての古人類学は、1856年のネアンデルタール人の発見から始まったが、初期の研究は1830年以来始まっていた。1859年までに現生人類と大型類人猿の形態的な類似性は議論されていたが、同年11月にチャールズ・ダーウィンが『種の起源』を著すまで「生物の進化」という概念は一般には正当化されなかった。ダーウィンの進化に関する最初の本は人類の進化についてはほとんど何も述べなかった。
- 「人類の起源と歴史に光が投げかけられるであろう。」
これがダーウィンが人類について述べた全てであった。それでも進化論の暗示は当時の読者にとって明らかだった。
トマス・ハクスリーとリチャード・オーウェンの論争は人類の進化に集中した。ハクスリーは1863年の著書『自然の中の人類の位置』で、類人猿と現生人類の多くの類似性と相違点について説得力を持って論じた。ダーウィンが『人間の由来と性選択』(1871)でその問題について論じる頃までにはその問題は広く知られ議論の的であった。チャールズ・ライエルとアルフレッド・ウォレスのようなダーウィンの支持者の多くも、現生人類の象徴的な精神性と道徳的な感性が自然選択によって形作られたという考えを好まなかった。
カール・フォン・リンネの頃から類人猿と現生人類は非常に似ているように見えるために、科学者たちは類人猿は人類の最も近い親類かもしれないと考えていた。19世紀にはゴリラ、チンパンジー、オランウータンのいずれが現生人類にもっとも近縁か論争があった。ダーウィンはチンパンジーかゴリラと考え、人類の祖先の化石が見つかるとしたらアフリカだろうと予測した。エルンスト・ヘッケルはオランウータンを人類にもっとも近縁と見なし、東南アジアから人類の祖先の化石が発見されるだろうと予測した。アフリカからは多くの化石人類が発掘された。一方ヘッケルの予測を信じたウジェーヌ・デュボワはインドネシアのジャワ島トリニールでジャワ原人の化石を発見し、後にこれがヒト属のホモ・エレクトゥスの亜種であるホモ・エレクトス・エレクトスに分類されている。

人類の祖先と思われる化石がアフリカで発見されたのはハクスリーやダーウィンの時代からしばらく後の1920年代であった。1925年にレイモンド・ダートはアウストラロピテクス・アフリカヌスを記載した。模式標本は洞穴の中から発掘されたアウストラロピテクスの幼児で、タウングの名にちなんでタウング・チャイルドと呼ばれた。南アフリカ共和国にある発見地タウングの洞窟では、コンクリートの原料が採掘されていた。タウング・チャイルドの化石は非常に保存状態の良い頭蓋骨を保持しており、頭蓋腔を推定できた。脳は小さかったが(410cm3)、その形は洗練されており、チンパンジーやゴリラのものよりも現代人に似ていた。また、化石は短い犬歯を持っており、大後頭孔の位置は直立二足歩行の証拠であった。これらの特徴全てはタウングチャイルドが二足歩行の人類の祖先で、類人猿から人類に変わりつつある証拠であるとダートに確信させた。しかしダートの主張は彼の発見に類似したより多くの化石が見つかるまで軽視され、真剣に検討されるまでに20年かかった。当時の主流な見解は二足歩行の前に脳の巨大化が起きたというものであり、現代人と同じような知性の発達が二足歩行の必要条件であると考えられていた。
アウストラロピテクスは現在、現生人類が属するヒト属の直接の祖先と考えられている。アウストラロピテクスとホモ・サピエンスは共にヒト亜族の一種である。しかし近年のデータは現生人類の直接の祖先としてアウストラロピテクス・アフリカヌスの位置に疑問を投げかける。彼らは進化的行き止まりの“従兄弟”であったかも知れない。アウストラロピテクスは当初、華奢なタイプと頑強なタイプに分類された。その後、頑強なアウストラロピテクスはパラントロプス属として分類し直されたが、一部の研究者はまだアウストラロピテクスの亜属と考えている。1930年に頑強なタイプが最初に記載されたとき、パラントロプス属が用いられた。1960年代に頑強な変種はアウストラロピテクスに加えられたが、近年では最初の分類どおり異なる属とする傾向がある。
ヒト属以前
|
真主齧上目 Euarchontoglires |
|
||||||||||||||||||||||||||||||

サル目以前





霊長類(サル目;霊長目)の進化の歴史は、約8500万年前まで遡ることができる。かつては有胎盤類の中で最も古い分類群と考えられていたが、現在は他の哺乳類も既にこの頃には分岐が進んでいたことが確認されている。霊長類は、同じく古い分類群で樹上生の祖先をもったであろうコウモリ類(翼手目)と共通祖先をもつと広く考えられていたが、解剖学的・形態学的、および、分子系統学的知見が発達するに連れて、ネズミ目(齧歯目)およびウサギ目と祖先を共有する一大分類群「真主齧上目」の一員と見なされるようになった。係る共通祖先の理論的に想定される生息年代は、白亜紀後期である。霊長類の既知で最古の化石は、白亜紀末期のララミディア大陸(元はローラシア大陸の一角、現在の北アメリカ大陸西部相当地域)で発見されており、プレシアダピス類(偽霊長類)と呼ばれている。このように、霊長類の進化は約6500万年前、白亜紀末期頃に始まったと考えられている。最初期の霊長類と考えられている動物は、白亜紀の北半球の陸地の大半を占めていながらも分裂する最中にあったローラシア大陸のどこかで誕生して拡散したと見られる。ローラシア大陸は中生代白亜紀から新生代初期にかけて分裂してゆくが、初期霊長類を載せた地域では北アメリカ大陸とヨーロッパ大陸に分かれてゆく。両大陸に分かれてゆく時代に初期霊長類は両方で繁栄し、約6550万年前から始まる新生代の暁新世と始新世の温暖な時代に、現生キツネザル類の祖先にあたるアダピス類 (Adapidae) と現生メガネザル類の祖先にあたるオモミス類 (Omomyidae. 真猿類の祖先) という二大グループを分岐させてから姿を消した。後述するが、その後、故郷ローラシアの後身にあたる北アメリカ大陸とヨーロッパ大陸は新生代氷河時代の本格的な到来で寒冷化し、霊長類はこれに耐え切れずに絶滅してしまう。命脈を保った霊長類は、北アメリカ大陸から姿を消す前にパナマ地峡を渡って南アメリカ大陸まで到達していたグループだけであったと考えられる。その後、アフリカ大陸にも霊長類が現れて隆盛するようになるが、ヨーロッパ経由で辿り着いたにしては生息年代に開きがありすぎるため、南アメリカ大陸から漂着したという説に説得力がある。なお、パナマ地峡の存在は霊長類の進化史において極めて重要で、アメリカ大陸全土における絶滅を防いだことも大きいが、北半球の霊長類を一掃した寒冷化の原因の一つにこの地峡の形成による海流の変化が考えられる。
サル目の特徴

霊長類(=サル目)は、次のような特徴を持つ。5本の指をもち、親指が他の4本と多少とも対向しているため、物をつかむことができる。前肢と後肢の指の爪は、ヒトを含めた狭鼻下目のすべての種ではすべての指の爪が平爪である。曲鼻猿亜目と広鼻下目の一部では平爪のほかに鉤爪をそなえる種もある。両目が顔の正面に位置しており、遠近感をとらえる立体視の能力に優れている。これらの特徴は、樹上生活において、正確に枝から枝に飛び移るために不可欠な能力である。多くの樹上性の哺乳類では、鉤爪を引っかけて木登りをするが、サル類の平爪はこれをあきらめ、代わりに指で捕まるか引っかかるかする方向を選んだものである。また、それが指先の器用さにつながることとなる。
直鼻猿亜目と曲鼻猿亜目の分岐
新生代に入り暁新世になるとアダピス類 (Adapidae) とオモミス類 (Omomyidae) が繁栄した。いずれもまだ原始的な種類で、アダピス類はのちの曲鼻猿類に、オモミス類はのちの直鼻猿類に進化したと考えられる。アダピス類とオモミス類はヨーロッパと北アメリカに分布したが、北アメリカの霊長類は寒冷化による森林の減少で絶滅し、旧世界を舞台に霊長類の進化は進んだ。曲鼻猿類の一部は海によって他の大陸から隔絶されていたマダガスカル島にアフリカから進出し(恐らくは流木等に掴まっての漂着)、キツネザル類に進化していった。
霊長類でL-グロノラクトンオキシダーゼ(ビタミンC合成酵素)の活性が失われたのは約6300万年前であり、サル目が直鼻猿亜目(酵素活性なし)と曲鼻猿亜目(酵素活性あり)との分岐が起こったのとほぼ同時である。ビタミンC合成能力を失った直鼻猿亜目にはメガネザル下目や真猿下目(サル、類人猿、ヒト)を含んでいる。ビタミンC合成能力を有する曲鼻猿亜目には、キツネザルなどが含まれる。なお、ビタミンC合成能力を失った動物は、ビタミンCを摂取しないとコラーゲンを合成できなくなり壊血病を発症して生存を維持できなくなる。直鼻猿亜目が遺伝子変異によりビタミンC合成能力を失ったにもかかわらず継続的に生存し得た最大の理由は、直鼻猿亜目が樹上生活で果物等のビタミンCを豊富に含む食餌を日常的に得られる環境にあったためである。
真猿下目とメガネザル下目の分岐
直鼻猿亜目は、その後、真猿下目とメガネザル下目に分岐する。この分岐の際に、真猿下目のX染色体に位置する錐体視物質に関連した色覚の多型が顕著になり、ヘテロ接合体の2本のX染色体を持つメスに限定した3色型色覚の再獲得につながり、さらに狭鼻下目のオスを含めた種全体の3色型色覚の再獲得へとつながる。
狭鼻下目と広鼻下目の分岐
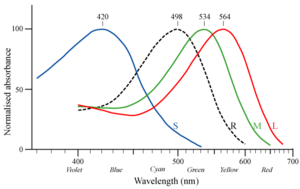
真猿下目が狭鼻下目(旧世界サル)と広鼻下目(新世界サル)に分岐したのは、3000-4000万年前と言われている。 脊椎動物の色覚は、網膜の中にどのタイプの錐体細胞を持つかによって決まる。魚類、両生類、爬虫類、鳥類には4タイプの錐体細胞(4色型色覚)を持つものが多い。よってこれらの生物は長波長域から短波長域である近紫外線までを認識できるものと考えられている。一方ほとんどの哺乳類は錐体細胞を2タイプ(2色型色覚)しか持たない。哺乳類の祖先である爬虫類は4タイプ全ての錐体細胞を持っていたが、2億2500万年前には、最初の哺乳類と言われるアデロバシレウスが生息し始め、初期の哺乳類は主に夜行性であったため、色覚は生存に必須ではなかった。結果、4タイプのうち2タイプの錐体細胞を失い、青を中心に感知するS錐体と赤を中心に感知するL錐体の2錐体のみを保有するに至った。これは赤と緑を十分に区別できないいわゆる「赤緑色盲」の状態である。この色覚が哺乳類の子孫に遺伝的に受け継がれることとなった。ヒトを含む旧世界の霊長類(狭鼻下目)の祖先は、約3000万年前、性染色体であるX染色体にL錐体から変異した緑を中心に感知する新たなタイプの錐体(M錐体)視物質の遺伝子が出現し、ヘテロ接合体の2本のX染色体を持つメスのみが3色型色覚を有するようになり、さらにヘテロ接合体のメスにおいて相同組換えによる遺伝子重複の変異を起こして同一のX染色体上に2タイプの錐体視物質の遺伝子が保持されることとなりX染色体を1本しか持たないオスも3色型色覚を有するようになった。これによって、狭鼻下目に第3の錐体細胞が「再生」された。3色型色覚はビタミンCを多く含む色鮮やかな果実等の発見と生存の維持に有利だったと考えられる。霊長類の3色型色覚の適応的意義については結論が出ていないのが現状である(上記「果実説」のほか,「若葉説」や「皮膚色説」も存在する。)。
なお、時代を下ってヒトの色覚に鑑みるに、ヒトが属する狭鼻下目のマカクザルに色盲がヒトよりも非常に少ないことを考慮すると、ヒトの祖先が狩猟生活をするようになり3色型色覚の優位性が低くなり、2色型色覚の淘汰圧が下がったと考えられる。色盲の出現頻度は狭鼻下目のカニクイザルで0.4%、チンパンジーで1.7%であり、現生のアフリカ系男性で2-4%、日本人男性で約5%、フランス、北欧系の男性で約10%である。広鼻下目のヨザル属は1色型色覚であり、ホエザル属は狭鼻下目と同様に3色型色覚を再獲得しているとされている。他方、ホエザルは一様な3色型色覚ではなく、高度な色覚多型であるとの指摘もある。これらのヨザル、ホエザルを除き残りの新世界ザル(広鼻下目)はヘテロ接合体のX染色体を2本持つメスのみが3色型色覚を有し、オスは全て色盲である。これは狭鼻下目のようなX染色体上での相同組換えによる遺伝子重複の変異を起こさなかったためである。ヒトは上記のような初期哺乳類と霊長目狭鼻下目の祖先のX染色体の遺伝子変異を受け継いでいるため、L錐体のみを保持したX染色体に関連する赤緑色盲が伴性劣性遺伝をする。男性ではX染色体の赤緑色盲の遺伝子を受け継いでいると色盲が発現し、女性では2本のX染色体とも赤緑色盲の遺伝子を受け継いでいる場合に赤緑色盲が発現する。なお、日本人では男性の5%、女性の0.2%が先天赤緑色覚異常であるとされる。
3000万年前、漸新世初期に現在の気候が始まると最初の南極の氷が形成され、アフリカと南アジア以外の霊長類は絶滅へ向かった。当時の霊長類の一つが曲鼻猿亜目キツネザル科に近いノタルクタスである。
生き残った熱帯の集団は(それらはカイロの南西ファイユーム低地の後期始新世と初期漸新世の化石層でよく見られる)現生の全霊長類を、すなわち曲鼻猿亜目に属するマダガスカルのキツネザル、東南アジアのロリス、アフリカのガラゴ、そして直鼻猿亜目に属する広鼻猿類(新世界ザル)と狭鼻猿類に属する旧世界ザル、大型類人猿、人類を生み出した。
新世界である南米の広鼻猿類(広鼻下目)は3000万年前から化石記録に現れるが、北アフリカの化石種で彼らの祖先に近縁なものは特定されていない。もしかすると西アフリカで異なる形態で生きていたのかも知れない。西アフリカからは、まだ解明されていない手段で南アメリカ大陸まで、霊長類、齧歯類、ボア、シクリッドが渡っている。洪水などで流されて大西洋経由で漂着したなどの可能性が考えられるも、決定的な説を見いだせていない。これに対して、広鼻下目(新世界サル)の祖先やテンジクネズミ上科の祖先がアフリカでできた浮島に乗って大西洋を流されて新世界の南アメリカ大陸に到着したという説も紹介されている。
既知のもっとも初期の狭鼻猿類は北ケニア地溝帯のEragaleitから見つかっているカモヤピテクス (Kamoyapithecus) で、2400万年前頃生きていたと見られている。その祖先は恐らく、エジプトピテクスかプロピリオピテクス (Propliopithecus) かパラピテクス (parapithecus) の近縁種と見られ、それらは3500万年前のファイユームの地層から見つかっている。その間の1100万年を繋ぐ化石は見つかっていない。
ヒト上科とオナガザル上科の分岐
霊長類の狭鼻下目がヒト上科とオナガザル上科に分岐したのは、3500万年前から3000万年前頃というのがゲノムベースの分析による2000年代前後の定説であったが、ミシガン大学の研究チームによる2010年発表の新説では、数百万年若い2800万年前頃から2400万年前頃と推定された。ヒト上科(テナガザル、オランウータン、チンパンジー、ゴリラ、ヒト)の共通の祖先が旧世界のサルから分枝した際に、尿酸オキシダーゼ活性が消失したものと推定される。尿酸オキシダーゼ活性の消失の意味付けは、尿酸が直鼻猿亜目で合成能が失われたビタミンCの抗酸化物質としての部分的な代用となるためである。しかし、ヒトを含むヒト上科では、尿酸オキシダーゼ活性の消失により難溶性物質である尿酸をより無害なアラントインに分解できなくなり、尿酸が体内に蓄積すると結晶化して関節に析出すると痛風発作を誘発することとなる。
テナガザルを含めた現生類人猿(=ヒト上科)では尾は失われている。
中新世初期にあたる約2200万年前、東アフリカの樹上棲に適応した初期の多種の狭鼻猿類は、それ以降の多様化のきっかけとなった。約2000万年前の化石は初期の旧世界ザルに属するビクトリアピテクス (Victoriapithecus) と思われる断片も含む。そのほかの形態は現生類人猿に近縁であるという明白な証拠はないが、類人猿に分類されている。現在認められているこのグループの属には、プロコンスル、ラングワピテクス (Rangwapithecus)、デンドロピテクス (Dendropithecus)、リムノピテクス (Limnopithecus)、ナコラピテクス、エクアトリウス (Equatorius)、ニャンザピテクス (Nyanzapithecus)、アフロピテクス (Afropithecus)、ヘリオピテクス、ケニアピテクス(ケニヤ…、ケニャ…、Kenyapithecus)がおり、全て東アフリカから1300万年以前に見つかっている。
1980年代にドイツで見つかった化石は約1650万年前のもので、東アフリカで発見された類似した化石よりも150万年ほど古いと考えられた。それは最初に大型類人猿の系統が現れたのがアフリカでなくユーラシアであったかも知れないと示唆する。約1700万年前にこの2つの大陸が地中海の拡大によって切り離される直前に、ヒト科の初期の祖先がアフリカからユーラシアへ渡ったのかも知れない。これらの霊長類がユーラシアで繁栄し、アフリカ類人猿と人類を生むことになる系統(ドリオピテクス)がヨーロッパまたは西アジアからアフリカに南下した。
遥かに離れた発掘地から中期中新世の旧世界ザルではない骨格が見つかっている。ナミビアの洞窟からオタビピテクス (Otavipithecus)、フランス、スペイン、オーストリアから、ピエロラピテクス (Pierolapithecus) とドリオピテクス (Dryopithecus) などである。それらは中新世初期から中期のアフリカと地中海沿岸が比較的暖かく穏やかな気候で、霊長類の多様化を促した証拠である。
中新世のヒト上科の証拠でもっとも新しいものはイタリアのオレオピテクス (Oreopithecus) で、約900万年前の石炭層から見つかっている。
ヒト上科の下位分類
ヒト上科からヒト亜族までの分類は、学説によって数種類に分かれる。以下は、オランウータン類をヒト科の下位に分類する学説に基づくものである。
表記は左から順に、1. 学名、2. あれば和名、( )内は別の和名や和名の表記揺れやその他の補足情報、3. 学名と表記の異なる英語名があればそれを含む全ての英語名、4. 特記事項。†(短剣符)は絶滅の意。incertae sedis は地位未確定の意。
- suprafamilia Hominoidea ヒト上科(ホミノイド) superfamily Hominoidae, hominoids
- †familia Dendropithecidae デンドロピテクス科 family Dendropithecidae
- †subfamilia incertae sedis (亜科不確定)
- †genus Dendropithecus デンドロピテクス属 genus Dendropithecus …2種。
- †genus Simiolus シミオルス属 genus Simiolus …5種。
- †subfamilia Nyanzapithecinae ニャンザピテクス亜科 subfamily Nyanzapithecinae
- †genus Nyanzapithecinus ニャンザピテクス属 genus Nyanzapithecinus …3種。
- †genus Mabokopithecus マボコピテクス属 genus Mabokopithecinus …1種。
- †genus Rangwapithecus ラングワピテクス属 genus Rangwapithecinus …1種。
- †genus Turkanapithecus トゥルカナピテクス属 genus Turkanapithecinus …1種。
- †genus Micropithecus (incertae sedis) ミクロピテクス属 genus Micropithecus …1種。
- †subfamilia incertae sedis (亜科不確定)
- †familia incertae sedis (科不確定)
- †subfamilia incertae sedis (亜科不確定)
- †tribus Dryopithecini ドリオピテクス族 tribus Dryopithecini
- †genus Kenyapithecus ケニアピテクス属(ケニヤ…、ケニャ…) genus Kenyapithecinus …1種。
- †genus Danuvius ダヌヴィウス属(ダヌビウス属) genus Danuvius …1種。
- †genus Ouranopithecus オウラノピテクス属 genus Ouranopithecus …2種。
- †genus Otavipithecus オタヴィピテクス属(オタビピテクス属) genus Otavipithecus …1種。
- †genus Oreopithecus オレオピテクス属 genus Oreopithecus …1種。
- †genus Nakalipithecus ナカリピテクス属 genus Nakalipithecus …1種。
- †genus Anoiapithecus アノイアピテクス属 genus Anoiapithecus …1種。
- †genus Dryopithecus ドリオピテクス属 genus Dryopithecus …2種。
- †genus Rudapithecus ルダピテクス属 genus Rudapithecus …1種。
- †genus Samburupithecus サンブルピテクス属 genus Samburupithecus …1種。
- †genus Udabnopithecus ウダブノピテクス属 genus Udabnopithecus …1種。
- †tribus Dryopithecini ドリオピテクス族 tribus Dryopithecini
- †subfamilia incertae sedis (亜科不確定)
-
familia Hylobatidae テナガザル科 family Hylobatidae, "lesser apes"
- genus Hylobates テナガザル属 genus Hylobates …7種。
- genus Hoolock フーロックテナガザル属 genus Hoolock …2種。
- genus Symphalangus フクロテナガザル属 genus Symphalangus …1種。
- genus Nomascus クロテナガザル属 genus Nomascus …6種。
- †familia Dendropithecidae デンドロピテクス科 family Dendropithecidae
(2017年版)

● Afropithecidae アフロピテクス科の枝
● Hylobatidae テナガザル科の枝
● Proconsulidae プロコンスル科(デンドロピテクス科)の枝
● Hominidae ヒト科の幹
● Ponginae オランウータン亜科の枝
● Lufengpithecini ルーフェンピテクス族の枝
● Sivapithecini シヴァピテクス族の枝
● Pongini オランウータン族の枝
● Homininae ヒト亜科の幹
● Dryopithecini ドリオピテクス族の枝
● Gorillini ゴリラ族の枝
● Hominini ヒト族の幹
● Pan チンパンジー属の枝
● Australopithecina アウストラロピテクス亜族の幹
● Homo ヒト属の幹
-
familia Hominidae ヒト科(ホミニド) family Hominidae, hominids, "great apes"
- †subfamilia incertae sedis (亜科不確定)
- †genus Pierolapithecus ピエロラピテクス属 genus Pierolapithecus …1種。
-
subfamilia Ponginae オランウータン亜科 subfamily Ponginae
- †tribus Lufengpithecini ルーフェンピテクス族(ルーフェングピテクス族) tribus Lufengpithecini
- †genus Lufengpithecus ルーフェンピテクス属(ルーフェングピテクス属) genus Lufengpithecus
- †Lufengpithecus lufengensis
- †Lufengpithecus keiyuanensis
- †Lufengpithecus hudienensis
- †genus Lufengpithecus ルーフェンピテクス属(ルーフェングピテクス属) genus Lufengpithecus
- †tribus Sivapithecini シヴァピテクス族 tribus Sivapithecini
- †genus Ankarapithecus アンカラピテクス属 genus Ankarapithecus
- †Ankarapithecus meteai
- †genus Sivapithecus シヴァピテクス属 genus Sivapithecus
- †Sivapithecus brevirostris
- †Sivapithecus punjabicus
- †Sivapithecus parvada
- †Sivapithecus sivalensis
- †Sivapithecus indicus
- †genus Gigantopithecus ギガントピテクス属 genus Gigantopithecus
- †genus Ankarapithecus アンカラピテクス属 genus Ankarapithecus
-
tribus Pongini オランウータン族(ポンゴ族) tribus Pongini
- †genus Khoratpithecus コラトピテクス属 genus Khoratpithecus
-
genus Pongo オランウータン属 genus Pongo, orangutans
- †Pongo hooijeri ベトナムオランウータン Vietnamese orangutan
- †Pongo weidenreichi
- Pongo abelii スマトラオランウータン Sumatran orangutan
- Pongo pygmaeus ボルネオオランウータン Bornean orangutan
- Pongo tapanuliensis タパヌリオランウータン Tapanuli orangutan
- †tribus Griphopithecini グリフォピテクス族 tribus Griphopithecini
- †genus Griphopithecus グリフォピテクス属 genus Griphopithecus
- †Griphopithecus alpani
- †Griphopithecus suessi
- †genus Griphopithecus グリフォピテクス属 genus Griphopithecus
- †tribus Lufengpithecini ルーフェンピテクス族(ルーフェングピテクス族) tribus Lufengpithecini
-
subfamilia Homininae ヒト亜科 subfamily Homininae
- (異説:†tribus Sivapithecini)…シヴァピテクス族をヒト亜科に分類する説ではこの位置に収まる。
-
tribus Gorillini ゴリラ族 tribus Gorillini
- †genus Chororapithecus コロラピテクス属 genus Chororapithecus
- †Chororapithecus abyssinicus コロラピテクス・アビシニクス(アビッシニクス)
-
genus Gorilla ゴリラ属 genus Gorilla, gorillas
-
Gorilla gorilla ニシゴリラ Western gorilla
- Gorilla gorilla gorilla ニシローランドゴリラ Western lowland gorilla
- Gorilla gorilla diehli クロスリバーゴリラ Cross River gorilla
-
Gorilla beringei ヒガシゴリラ Eastern gorilla
- Gorilla beringei beringei マウンテンゴリラ Mountain gorilla
- Gorilla beringei graueri ヒガシローランドゴリラ(グラウアーゴリラ) Eastern lowland gorilla, Grauer's gorilla
-
Gorilla gorilla ニシゴリラ Western gorilla
- †genus Chororapithecus コロラピテクス属 genus Chororapithecus
-
tribus Hominini ヒト族 tribus Hominini, hominins
-
subtribus Panina チンパンジー亜族 subtribus Panina
-
genus Pan チンパンジー属 genus Pan, chimpanzees
- Pan troglodytes チンパンジー(ナミチンパンジー) Common Chimpanzee
- Pan paniscus ボノボ(ピグミーチンパンジー) Bonobo
-
genus Pan チンパンジー属 genus Pan, chimpanzees
- (異説:subtribus Australopithecina アウストラロピテクス亜族 subtribus Australopithecina, Australopithecine, Australopithecines) …ヒト属に先行するヒト亜族的分類群を独立した亜族と見なす学説における地位。
- subtribus Hominina ヒト亜族 subtribus Hominina …これより下位は「ヒト亜族の下位分類」を参照のこと。
-
subtribus Panina チンパンジー亜族 subtribus Panina
- †subfamilia incertae sedis (亜科不確定)
-
familia Hominidae ヒト科(ホミニド) family Hominidae, hominids, "great apes"

ヒト科とテナガザル科の分岐
分子的な証拠は、2000万年から1600万年前にヒト上科がヒト科とテナガザル科に分岐したことを示している。テナガザルの祖先を明らかにする化石史料は見つかっていない。彼らは東南アジアの未知のヒト科の集団から分かれたかも知れない。
ヒト亜科とオランウータン亜科の分岐
その後、ヒト科が1400万年前にヒト亜科とオランウータン亜科に分岐したと推定されている。初期のオランウータンは1000万年前のインドのラマピテクス、あるいはトルコのグリフォピテクスかもしれない。
ヒト族とゴリラ族の分岐
約1000万年前にヒト亜科がヒト族とゴリラ族に分岐したと推定されている。 ゴリラ、チンパンジー、ヒトを結び付ける最後の祖先はケニアで見つかったナカリピテクス、あるいはギリシャで見つかったオウラノピテクスの可能性が示唆されている。
ヒト亜族とチンパンジー亜族の分岐
約700万年前にヒト亜族とチンパンジー亜族に分岐したと推定されている。DNAの変異にかかる時間に基づき推定すると800-700万年前に分岐した可能性が高いとの論文が発表されている。ヒトのDNAはチンパンジーのDNAと98.4%同一である。ゴリラとチンパンジーの系統の化石は非常に限定的である。保存に厳しい環境(熱帯雨林土は酸性で、骨を分解しやすい)とサンプルの偏りがこの問題の原因である。彼ら以外のヒト科は赤道の外縁あたりで、アンテロープ、ハイエナ、ウマ、ゾウたちと共に、より乾燥した環境に適応した可能性がある。彼らの化石は比較的有名である。チンパンジー亜族と分岐し直立二足歩行をしていたヒト亜族のうち、もっとも初期のものはサヘラントロプス・チャデンシス(700-600万年前)である。
ヒト(=人類)とは、広義にはチンパンジー亜族と分岐したヒト亜族に属する動物の総称であり、狭義には現生の(つまり現在生きている)人類を指す。
ヒト亜族の下位分類
表記は左から順に、1. 学名、2. あれば和名、( )内は別名など補足情報、3. 学名と表記の異なる英語名があれば全ての英語名、4. 特記事項。†(短剣符)は絶滅の意。
- ヒト属に先行するヒト亜族的分類群を独立した亜族と見なす学説では、それらを束ねて以下の地位を与える。
- †subtribus Australopithecina アウストラロピテクス亜族 subtribus Australopithecina, Australopithecine, Australopithecines
- (異説:†genus Graecopithecus グレコピテクス属(グラエコピテクス属) genus Graecopithecus …約720万年前。オウラノピテクス属 (Ouranopithecus) のシノニム。
- †genus Sahelanthropus サヘラントロプス属 genus Sahelanthropus …約700万年前。
- †Sahelanthropus tchadensis サヘラントロプス・チャデンシス …発見された1個体は「トゥーマイ猿人(英: Toumaï)」の愛称をもつ。
- †genus Orrorin オロリン属 genus Orrorin …約610万年~約580万年前。
- †genus Ardipithecus アルディピテクス属 genus Ardipithecus …約580万年~約440万年前。
- †Ardipithecus kadabba アルディピテクス・カダッバ(カダッバ猿人)
- †Ardipithecus ramidus アルディピテクス・ラミドゥス(ラミドゥス猿人)
- †genus Australopithecus アウストラロピテクス属(オーストラロピテクス属 以下同様) genus Australopithecus …旧称“華奢型アウストラロピテクス”。約540万年~約150万年前。
- †Australopithecus afarensis アウストラロピテクス・アファレンシス(アファール猿人)
- †Australopithecus africanus アウストラロピテクス・アフリカヌス(アフリカヌス猿人)
- †Australopithecus anamensis アウストラロピテクス・アナメンシス(アナム猿人)
- †Australopithecus bahrelghazali アウストラロピテクス・バーレルガザリ(バーレルガザリ猿人)
- †Australopithecus garhi アウストラロピテクス・ガルヒ(ガルヒ猿人)
- †genus Kenyanthropus ケニアントロプス属(ケニヤン…、ケニャン…) genus Kenyanthropus …約300万年~約270万年前。
- †genus Paranthropus パラントロプス属 genus Paranthropus …旧称“頑丈型アウストラロピテクス”。約270万年~約120万年。
- †Paranthropus aethiopicus パラントロプス・エチオピクス(エチオピクス猿人)
- †Paranthropus robustus パラントロプス・ロブストゥス(ロブストゥス猿人)
- †Paranthropus boisei パラントロプス・ボイセイ(ボイセイ猿人)
- genus Homo ホモ属(ヒト属) genus Homo, humans …約250万年前~現世。これより下位は「ヒト属の下位分類」を参照のこと。
アウストラロピテクス
アウストラロピテクス(学名:Australopithecus、和名の表記揺れ:オーストラロピテクス)は、アフリカで生まれた初期の人類であり、約400万年前 - 約200万年前に生存していた、いわゆる華奢型の猿人である。身長は120cm台 - 140cm台くらいで、脳容積は現生人類の約35%の500cc程度であり、チンパンジーとほとんど変わらないが、骨格から二足歩行で直立して歩く能力を持つと考えられている。アウストラロピテクス・アフリカヌスの頭蓋骨には人類と同じ直立二足歩行の姿勢であったことを示す位置に脊柱とつながる穴(大後頭孔)があったことからである。姿形は直立したチンパンジーというイメージである。以前は最も古い人類の祖先とされていたがアルディピテクス属の発見により、その次に続く属となった。約440万 - 約390万年前にA・アナメンシスが、約390万 - 約300万年前にアファレンシスが現れ、約330万 - 約240万年前にアウストラロピテクス・アフリカヌスに進化した。この属からパラントロプスと、ホモ(ヒト属)最初の種ホモ・ハビリスに進化したと考えられている。
パラントロプス
パラントロプス(学名:Paranthropus)の体長は1.3から1.4mで、華奢型アウストラロピテクスよりひと回り大きい。脳もいくらか大きめである。 形態的には、アウストラロピテクスよりヒト的な特徴は減少しており、堅い食物を咀嚼するため、高く厚い下顎と太い側頭筋、それを通すために張り出した頬骨弓および大型の臼歯など頑丈な咀嚼器を有している。硬い植物性の食物、根などを常食としていたと考えられる。
ヒト属
ヒト属(別名:ホモ属、学名:Genus Homo)は、ヒト亜族のうちで、大脳が著しく増大進化したタクソン(分類群)を指している。属名 Homo は、イタリック祖語起源のラテン語 homō(日本語音写例:ホモー、英訳:a human being , a man, a person、和訳:人間、人)に由来し、「分類学の父」と称される博物学者カール・フォン・リンネが、生きとし生けるものを初めて分類するにあたって自分達の学名として選んだ名称である。

ただし、ヒト属の分散と類縁関係について統一的見解は存在しない(以下同様)。

現代の分類学において、"解剖学的現代人"の意味合いをもつ「現生人類」は、学名でいう Homo sapiens(亜種を認める説では Homo sapiens sapiens)と、結果的の同義であり、ヒト属で唯一の現存するタクソンであるからこそ、その名で呼ばれている。現生人類の起源についての研究が進むに連れ、ほかにもヒト属の下位タクソンが存在したものの、現生人類という1タクソンを残してそれ以外全て絶滅していることが分かっている。絶滅グループの中に現生人類の直接の祖先が(それが既知の種か未発見種かはともかくも)いることに疑問の余地は無いが、絶滅グループのほとんどは現生人類の"従兄弟"であって、ヒト属の範疇にある化石人類と現生人類が形成している進化系統群のなかでの個々の種の分類学上の地位(進化上の位置、リンネ式分類法上の階級など)について統一的見解が得られたことはない。これは化石人類の分類に用いられる種の概念が解剖学的特徴に基づいた形態的種であるためであり、2つの種の中間的な特徴をもつがゆえに分類の困難な化石も多く発見されている。「種 (分類学)」も参照のこと。
サハラ砂漠の拡張が初期のヒト属の進化の原因となったともいわれているが、ヒト属の進化の要因についていくつかの説がある。一つの説はサバンナ説で、人類学者レイモンド・ダートによって提示された。樹上性であった(かもしれない)人類の祖先の一部が、乾燥化に伴う樹林の減少によってやむを得ず、あるいは、繁栄したがための生息域の積極的拡大もしくは弱小グループの追放という形で、新天地を求めてサバンナへ進出したというものである。もう一つはアクア説と呼ばれており、こちらには多くの研究者が異論を唱えている。これは食料を集めるために水中を歩き、泳ぎ、潜ることが人類の祖先と他の類人猿の祖先に異なる選択圧を与えたと主張している。フランスの古人類学者イヴ・コパンは東アフリカの大地溝帯が引き起こした東側地域の乾燥化が、チンパンジー属とヒト属の祖先グループを西側の森と東側のサバンナに分断し、それぞれが地理的種分化によって別属となったという仮説(イーストサイドストーリーと呼ばれる)を提唱したが、大地溝帯の西側からも祖先系統と見られる化石種が発見されたことで、2003年に提唱者自らこの仮説を撤回している。
考古学と古生物学の証拠に基づいて、さまざまなヒト属の食性を推論することが可能で、食性がヒト属の身体と行動に与えた進化的影響は研究の中途にある。
現生しているヒトの脳が肥大化・高度化した原因として推測されている遺伝子変異はいくつか挙げられている、一部を挙げるとASPM、CMAH、DUF1220、FOXP2、HAR1、HARE5、MCPH、等がある。
ヒト属の下位分類
表記は左から順に、1. 学名、2. あれば和名、( )内は別名など補足情報、3. 学名と表記の異なる英語名があれば全ての英語名、4. 特記事項。†(短剣符)は絶滅の意。
-
genus Homo ホモ属(ヒト属) genus Homo …約250万年前~現世。これより上位は「ヒト亜族の下位分類」を参照のこと。
- †Homo habilis ホモ・ハビリス(ハビリス原人)
- †Homo rudolfensis ホモ・ルドルフエンシス(ホモ・ルドルフェンシス)
- †Homo ergaster ホモ・エルガステル(ホモ・エルガスター) Homo ergaster, Homo erectus ergaster, African Homo erectus
- †Homo erectus ホモ・エレクトゥス(…エレクトス 以下同様)
- †Homo erectus erectus ホモ・エレクトゥス・エレクトゥス(ジャワ原人) Java man
- †Homo erectus pekinensis ホモ・エレクトゥス・ペキネンシス(北京原人) Peking man
- †Homo erectus mauritanicus ホモ・エレクトゥス・マウリタニクス(異説:ホモ・マウリタニクス。テルニフィヌ人) Ternifine man
- †Homo erectus yuanmouensis ホモ・エレクトゥス・ユァンモウエンシス(元謀原人) Yuanmou man
- †Homo erectus soloensis ホモ・エレクトゥス・ソロエンシス(ソロ人) Solo man
- †Homo rhodesiensis ホモ・ローデシエンシス(ローデシア人) Rhodesian man, Kabwe man
- †Homo cepranensis ホモ・ケプラネンシス(ホモ・セプラネンシス) Ceprano man
- †Homo georgicus ホモ・ゲオルギクス(ドマニシ人、ドマニシ原人) Dmanisi man
- †(onbepaald; 未定義・未記載) 馬鹿洞人 Red Deer Cave people, Edelhert-grotmense …2020年の知見で、全ての化石人類のなかで最後まで生き残っていたグループ。
- †Homo floresiensis ホモ・フローレシエンシス(フローレス人、フローレス原人) Flores man, "hobbit" …エレクトゥス系統群の傍流で、島嶼化している。
- †Homo naledi ホモ・ナレディ …エレクトゥス系統群の傍流で、矮小化している。
- †Homo antecessor ホモ・アンテセッサー(…アンテセッソール、…アンテケッソール) …エレクトゥス系統群の傍流ではあるが、本流(ハイデルベルゲンシスに到る進化系統)に近い。
- †Homo heidelbergensis ホモ・ハイデルベルゲンシス(ハイデルベルク人) Heidelberg man …エレクトゥス系統群からの分枝で、サピエンス種およびネアンデルターレンシス種の共通祖先もしくはその近縁種。
- †Homo neanderthalensis ホモ・ネアンデルターレンシス(ネアンデルタール人、古典的ネアンデルタール、ネアンデルタール人類) Neanderthal man …サピエンス種と混血した可能性あり。
- †(onbepaald; 未定義・未記載:Homo sapiens ssp. Denisova, Homo sapiens ssp. Altai ) デニソワ人 Denisovans, Denisova hominins, Denisova-hominien, Denisova people …サピエンス種と混血した可能性あり。
-
Homo sapiens ホモ・サピエンス(ヒト)
- †Homo sapiens idaltu ホモ・サピエンス・イダルトゥ(…イダルツ、ヘルト人) Herto man
- Homo sapiens sapiens ホモ・サピエンス・サピエンス(現生人類、解剖学的現代人) anatomically modern humans, modern humans
原人
ホモ・ハビリス
ホモ・ハビリス(学名:Homo habilis)は、およそ240万から140万年前にかけて生存していた。ヒト属の最初の種であるハビリスは鮮新世後期か更新世初期に南アフリカと東アフリカで出現した。おそらく250万から200万年前にアウストラロピテクスの一つから種分化したと考えられている。ハビリスはアウストラロピテクスよりも小さな臼歯と大きな脳を持っており、石と、おそらく動物の骨から道具を製造した。彼らは初めて知られたヒト科の一種で、発見者ルイス・リーキーによって彼らの石器に結び付けて「器用な人」とあだ名を付けられた。一部の科学者は頭蓋後部の形態からホモ・サピエンスのような二足歩行よりも樹上に適応していたと考え、ヒト属からアウストラロピテクス属へ移すよう提案している。
ホモ・ルドルフエンシスとホモ・ゲオルギクス
これらは約190万年から約160万年前の化石に名付けられた種である。ホモ・ハビリスとの類縁関係は明白ではない。
ホモ・ルドルフエンシス(学名:Homo rudolfensis、和名の表記揺れ:ホモ・ルドルフェンシス)は、ケニアで発見された一つの不完全な頭骨である。研究者はハビリスの一種であると主張したが、まだ確かめることができない。
ホモ・ゲオルギクス(学名:Homo georgicus)は、ジョージアで発見された。ホモ・ハビリスとホモ・エレクトゥスの中間か、あるいはホモ・エレクトゥスの亜種であるかも知れない。
ホモ・エルガステルとホモ・エレクトゥス

ホモ・エレクトゥス(学名:Homo erectus、和名の表記揺れ:ホモ・エレクトス)の最初の化石は、1891年にインドネシアのジャワ島でオランダ人軍医ウジェーヌ・デュボワによって発見された。彼は当初、その化石が人類と類人猿の中間であると考え、ピテカントロプス・エレクトゥスの名を与えた。ホモ・エレクトゥスは約180万年前から約7万年前までという非常に長い期間に亘って生存していた。約150万年~約100万年前、更新世初期に脳がより大きくなり精巧な道具を作ったホモ・ハビリスの子孫がアフリカ、アジア、ヨーロッパの各地に分散した。これらの特徴は古人類学者にとって彼らをホモ・ハビリスとは異なる種に分類するのに十分な理由となる。しばしば初期の段階、180万から125万年前までは別の種ホモ・エルガステル、あるいはエレクトゥスの亜種ホモ・エレクトゥス・エルガステルと扱われることがある。
エレクトゥスは間違いなく直立二足歩行していた事が明らかな最初の人類の祖先で、それはしっかり嵌まる膝蓋骨と大後頭孔(脊椎が入る頭骨の孔)の位置の変化によって可能になった。加えて彼らは肉を調理するために火を使った可能性がある。ホモ・エレクトゥスの有名な例は北京原人である。多くの古人類学者はホモ・エルガステルという呼称をこのグループの非アジア種に用いていて、エレクトゥスと言う呼称はアジア地域で見つかり、エルガステルとわずかに異なる骨格、歯の特徴を満たしている化石だけに用いているが、本項ではその用法に従っていない。
ホモ・ケプラネンシスとホモ・アンテセッサー
これらはホモ・エレクトゥスとホモ・ハイデルベルゲンシスの間をつなぐかも知れないと主張されている。
ホモ・アンテセッサー(学名:Homo antecessor、和名の表記揺れ:ホモ・アンテセッソール、ホモ・アンテケッソール)は、約120万年前から約50万年前にかけて生存していた。スペインとイングランドから化石が発見されている。
ホモ・ケプラネンシス(学名:Homo cepranensis、和名の表記揺れ:ホモ・セプラネンシス)は、イタリア半島から一つの頭骨片として発見されている。約80万年前のものと推測されている。
ホモ・ハイデルベルゲンシス
ホモ・ハイデルベルゲンシス(学名:Homo heidelbergensis、別の和名:ハイデルベルク人)は、約80万年から約30万年前にかけて生存していたタクソンである。
より高度な進化を遂げた人類としてホモ・エレクトゥスの系統群から分岐したタクソンであるが、階級的位置づけについては大きく分けて2つの見解がある。一つは原人の進化段階を脱していないという捉え方で、これを反映して提案された学名は Homo erectus heidelbergensis(ホモ・エレクトゥス・ハイデルベルゲンシス)である。次に、過渡期にあることを重視して提案された学名 Homo heidelbergensis(ホモ・ハイデルベルゲンシス)があり、これが多くの支持を集めている。これらに加えてハンガリーの人類学者アンドール・トマが後頭骨化石標本サムを論拠として唱えた仮称 Homo erectus seu sapiens paleohungaricus(ホモ・エレクトゥス・セウ・サピエンス・パレオフンガリクス。意訳:ホモ・エレクトゥス・パレオフンガリクス、もしくは、ホモ・サピエンス・パレオフンガリクス)もあるが、支持者が多いとは言い難い。
現生人類へ到る進化の道筋を"本流"とするなら、まさにそれ以前に本流を内包していたエレクトゥス系統群から分岐し、次の時代の本流そのもの、あるいは本流への繋ぎ役になったと考えられている。
ホモ・フローレシエンシス

ホモ・フローレシエンシス(学名:Homo floresiensis、別の和名:フローレス人)は、約19万年から約5万年前という比較的最近を生存期間とするタクソンである(※発見当初は約10万年~約1.2万年前と推定された)。彼らの体格は極めて小さく、これは島嶼化によるものと考えられている。「ホビット」という愛称もこれに由来する。
フローレシエンシス種の主要な発見は、2003年、ニュージーランドの考古学者マイク・モーウッドによってインドネシアのフローレス島から発見された30歳前後と思しき女性の骨格化石であり、彼女の生存年代は約1.8万年前と見積もられた(※この数値は今では支持を失っている)。この女性の推定される生前の姿は、身長わずか1メートルで、脳容量は380cm3とチンパンジー並みに小さく、現代人女性(平均1400cm3)の3分の1程度でしかなかった。
フローレシエンシス種は、その矮小さと年齢から、実際に最近まで生きていた現生人類と共通しない特徴をもつホモ属の興味深い例と考えられている。すなわち、いつの時点かで現生人類と祖先を共有するが、現生人類の系統群とは異なる独自の進化過程を辿ったと思われ、多くの研究者はエレクトゥスの系統群に共通祖先が含まれると考えている。しかし、彼らが本当に別の種であるかは未だ議論が続いており、小人症を患った現生人類と考える研究者も一部にはいる。この仮説は、フローレス島に住む現代人が小柄であるために、ある程度説得力がある。小柄さと小人症によって本当にホビットのような矮小な地域個体群が生まれた可能性はある。別種説への他の主要な反論は、現生人類と関連した道具類とともに発見されたという点である。
しかしいずれにしても、現在知り得る進化系統樹を結果から見れば、彼らの枝の先には何も生まれなかった。その意味で、進化上の完全な傍流で終わった。
旧人
ホモ・ヘルメイ
ホモ・ヘルメイ(仮称:Homo helmei)は、約26万年前 (259±35 ka) に生きていたと推定されるフロリスバッド人 (1932年出土。cf. en:Florisbad Skull) に与えるべく提案されている仮の学名の一つ。パラントロプスあたりから分岐して独自に進化してきた原人と見なす学説の下では、Africanthropus helmei(アフリカントロプス・ヘルメイ)の学名が提案されている。しかし、原人ホモ・ハイデルベルゲンシスに含まれるとする説が有力であり、この説の下で提案されている学名は、当然ながら、ホモ・ハイデルベルゲンシスと同じである。また、ホモ・ハイデルベルゲンシスから分岐したタクソンと見なす説もあり、この場合に Homo helmei(ホモ・ヘルメイ)という学名が成立し得る。このヘルメイ種は、古代型ホモ・サピエンス(早期ホモ・サピエンス;en:early Homo sapiens)の一種である可能性が唱えられている。
ホモ・ローデシエンシス


ホモ・ローデシエンシス(学名:Homo rhodesiensis、別の和名:ローデシア人)は、約30万年前から約12.5万年前までの間、アフリカの比較的広範囲地域(南アフリカ、東アフリカ、北アフリカ)に生息していたと考えられている。
現生人類(ホモ・サピエンス)の古拙(アルカイック)形の一種、すなわち原始的1亜種と見なして、Homo sapiens rhodesiensis(ホモ・サピエンス・ローデシエンシス)という学名と、"Archaic Homo sapiens(アルカイック・ホモ・サピエンス)" の一種という英語名が提案されもしたが、多くの研究者は、本種をホモ・ハイデルベルゲンシスと同じか極めて近い類縁種と考えている。原人よりは進化し、現生人類よりは原始的であるため、旧人段階にあるという見解もある。一時期はホモ・ネアンデルターレンシスの1亜種と考えられることもあったが、現在ではそれとは種レベルで異なる旧人と見なされている。
- ガーウィーズの頭蓋骨
2006年2月16日、エチオピアはアファール盆地の一角を占めるガーウィーズ川流域 (Gawis river basin) にあるガーウィーズ (Gawis) にて、おそらくホモ・エレクトゥスとホモ・サピエンスの中間か、それに近い進化的傍流(行き止まり)に属する種のものと思われる頭蓋骨上部の化石が発見された。「ガーウィーズの頭蓋骨(ガーウィーズ・クレイニアム、- クレニアム)(en)」と呼ばれるこの化石標本は、層位学的には約50万年から約20万年前に属するものと推定された。推定された生息年代と生息地は、ホモ・ローデシエンシスと重なるところが多い。ただ、知られているのは概要のみで、発掘チームは査読付き論文を発表していない。標本の特徴は、彼らが中間種であるか、ボド・マンの女性のものであるかを示している。
ホモ・ネアンデルターレンシス

ホモ・ネアンデルターレンシス(学名:Homo neanderthalensis、別の和名:ネアンデルタール人、異称:古典的ネアンデルタール、ネアンデルタール人類)は、 約40万年前に出現し、2万数千年前に絶滅したとみられる。ただし、新しい学説(2014年発表)は約4万年前に絶滅したとする。
ネアンデルタール人は、ホモ属の独立種ホモ・ネアンデルターレンシスと見なすのが今日の世界の定説であるが、未だ発見されていない理論上のサピエンス種の原初的グループから早期に分岐した1亜種ホモ・サピエンス・ネアンデルターレンシスと見なす説が一部にはある(cf. 学名と異説)。既知で最古のサピエンス種であるイダルトゥの出現時期は約16万年前であるから、それより大きく先行していることになる。
ネアンデルタール人と現生人類(ホモ・サピエンス)の間での大規模な遺伝子流動、すなわち「混血」の可能性については、マックス・プランク進化人類学研究所所属の人類学者マーク・ストーンキングが、ペンシルベニア州立大学所属時代の1997年に「これらの(ネアンデルタール人の骨から抽出されたミトコンドリアDNAに基づく)結果は、ネアンデルタール人がミトコンドリアDNAを現代人に与えなかったことを示している。(...略...)ネアンデルタール人は我々の祖先ではない。」と語っており、ネアンデルタール人のDNAの塩基配列研究もこの結果を支持した。また、多地域進化説の支持者は最近の非アフリカ人の核DNAが100万年前まで遡る可能性を示すことを研究したが、現在この研究の信頼性は疑われている。古植物学者で地球生物学者 (geobiologist) のミヒャエル・クリングス (Michael Krings) らが2008年に発表したミトコンドリアDNA解析に基づく分子系統学的知見は、大規模な混血が起こらなかったことを示すとともに、両者が約66万年前という一層遠い昔に共通祖先をもつことをも示唆した(cf. 現代人との混血)。ところが、2010年になると混血の痕跡があるとする研究結果がによって『サイエンス』誌上で発表され、議論が収束する様子は見られない。
絶滅については、アメリカの人類学者ナオミ・クレッグホーン (Naomi E. Cleghorn. テキサス大学アーリントン校所属) は、イタリア半島やコーカサス山脈で約4万年前に相次いだ火山噴火を理由に挙げている。このような環境的要因を指摘する説は以前にも発表されていたが、約4万年前に起こった事象はその種の災害とは規模が違っており、例えば、複数の火山がほぼ同時期に噴火する苛烈なものであったという。なかでもヴェスヴィオ山周辺地域で約3万9000年前に発生したプリニー式噴火であるカンパニアン・イグニンブライト噴火は、ヨーロッパ大陸における過去20万年間で最も規模の大きい噴火であった。「当時のヨーロッパ大陸には現生人類の小集団も住んでいたので、噴火の影響を同様に受けたと考えられる。しかしながら、ネアンデルタール人のほとんどがヨーロッパに居住していたのに対し、現生人類はアフリカやアジアにより大きな人口を抱えていたため、絶滅を避けられたようだ。」とクレッグホーンは考える。
ネアンデルタール人は約3万年前に滅亡したと長らく考えられていたが、2005年、イベリア半島南端にある「ジブラルタルの岩」に属するゴーラム洞窟が化石人類の遺跡であると判明し、ネアンデルタール人に特有の石器類や、洞内で火を利用していた痕跡(炭化した松かさ〈イタリアカサマツの球果〉や焚火跡)が見つかったことで、細々にではあってももっと若い時代にまで命脈を保っていたことが分かった。これらの遺物のうち最も古いものは約12万5000年前に属し、そして、最も若いものは約2万8000年~約2万4000年前に属しているという、放射性炭素年代測定の数値が出たからである。洞窟の奥には約2万300年~約1万9500年前のものと推定されるネアンデルタール人以外の人類の手形が遺されており、住人の入れ替わりがあったことを確認できる。ゴーラム洞窟遺跡が真に最も若い時期のネアンデルタール人の痕跡であるなら、かつて豊かな草原の只中にあったのが厳しい半乾燥地帯に変わってしまい、いつしか侵蝕を受けてアルボラン海に面してしまった地域と考えられるヨーロッパ大陸南西端部の小さな洞窟で、彼らは最期を迎えたことになる。しかし当節の冒頭で述べたように、新たな学説は大きく異なる時期を示しており、統一的見解を得るには程遠い。
新人
ホモ・サピエンス・イダルトゥ

ホモ・サピエンス・イダルトゥ(学名:Homo sapiens idaltu、和名の表記揺れ:ホモ・サピエンス・イダルツ、別の和名:ヘルト人)はエチオピアで発見されており、16万年前頃生きていたと考えられる。それは亜種として扱われてはいるが(ただし、ホモ・サピエンスの亜種分類法については学説上統一した合意はない)、解剖学的には現代人であり、知られているなかで最も古い新人段階の人類である。彼らの直接の子孫がネグロイドであり、モンゴロイドとコーカソイドはネアンデルターレンシスとの混血種であるらしいという最近の研究結果がある。これによると、イダルトゥは系統的にネグロイドに属することになる。
イダルトゥよりさらに古いサピエンスの直接の祖先としては約26万年前のフロリスバッド人 (1932年出土。cf. en:Florisbad Skull) や金牛山人の人骨が発見されているが、これらは進化段階としては旧人とみられる。ただし、イスラエルで約40万年前の最古のホモ・サピエンスである可能性がある人骨が発見されている。ネアンデルタール人との共通祖先との分岐年代が40万年以上前であることから、分岐直後の時期にはホモ・サピエンスが存在していたという解釈も可能であり、その場合、上記の人骨化石はイダルトゥよりさらに古いホモ・サピエンスの発見ということになる。
ホモ・サピエンス


ホモ・サピエンス(学名:Homo sapiens)は、解剖学的に何ら違いが認められない現代の人類と化石人類とを同一の種と認めたうえでの、1タクソン(1分類群)である。“解剖学的現代人”の意味合いをもって呼ばれる「現生人類」と、同じものを指す。ラテン語 sapiens は「wise、賢い」の意。約25万年前に出現したと考えられ、その後、唯一の残存種となった。
現代人と上記のイダルトゥには亜種レベルの相違があるとみなして、亜種「Homo sapiens sapiens〈ホモ・サピエンス・サピエンス〉」として扱うこともあるが、ホモ・サピエンスの亜種については統一した合意はないため、本項目は「ホモ・サピエンス」とする。 47万年〜66万年前に上記ネアンデルタール人との共通祖先から古代型サピエンスが分岐した。ここでは旧人時代の古代型サピエンスについても記述する。40万年前から25万年前の中期更新世の第二間氷期までの間に、旧人段階であった彼らが頭骨の拡張と石器技術が発達したようで、この事がホモ・エレクトゥスからホモ・サピエンスへ移行の証拠と見られている。移行を示す直接の証拠は、ホモ・エレクトゥスがアフリカから他の地域へ移住した間にアフリカで種分化が起きたことで(アフリカのどこで起きたかについてはわかっていない)エレクトゥスからホモ・サピエンスが分かれたことを示唆している。その後アフリカとアジア、ヨーロッパでエレクトゥスがホモ・サピエンスに入れ替わった。このホモ・サピエンスの移動と誕生のシナリオは単一起源説(アフリカ単一起源説)と呼ばれていて、現在古人類学において多地域進化説と単一説で激しい議論がされている。また、人類の遺伝的多様性が他の種に比べると非常に小さいことを確認されているが、これは比較的最近に各地に分散したか、トバ山噴火の影響の可能性がある。
約7万5000年前から約7万年前に、インドネシア、スマトラ島にあるトバ火山が大噴火を起こして気候の寒冷化を引き起こし、その後の人類の進化に大きな影響を与えた。トバ・カタストロフ理論によれば、大気中に巻き上げられた大量の火山灰が日光を遮断して火山の冬を引き起こし、地球の気温は平均5℃も低下したという。劇的な寒冷化はおよそ6000年間続いたとされる。その後も気候は断続的に寒冷化するようになり、地球は最終氷期へと突入する。この時期まで生存していたホモ属の進化的源流にあたるタクソン(ホモ・エレクトゥスなど)の主要なグループは絶滅したと考えられる。トバ事変の後まで生き残ったホモ属はネアンデルタール人と現生人類のみである。現生人類も、トバ事変の気候変動によって総人口が1万人までに激減したという。かろうじて生き残った現生人類も人口減少によってボトルネック効果が生じ、その遺伝的多様性は失われた。現在、人類の総人口は70億人にも達するが、遺伝学的に見て、現生人類の個体数のわりに遺伝的特徴が均質であるのは、トバ事変のボトルネック効果による影響であるとされる。遺伝子の解析によれば、現生人類は極めて少ない人口(1000組~1万組ほどの夫婦)から進化したことが想定されている。遺伝子変化の平均速度から推定された人口の極小時期はトバ事変の時期と一致する。この学説は約6万年前に生きていた“Y染色体アダム”や約14万年前に生きていた“ミトコンドリア・イヴ”を想定した学説とは矛盾しない。また、現生人類の各系統が約200万年〜約6万年前の時期に分岐したことを示している現生人類の遺伝子の解析の結果も、トバ・カタストロフ理論とは矛盾しない。なぜならば、トバ・カタスロトフ理論は総人口が数組の夫婦まで減少したという学説ではなく、そこまで凄まじいボトル・ネック現象を想定しているわけではないからである。現生人類の遺伝的多様性は、トバ事変によって現生人類の人口が一度減少したことを示唆している。
また、衣服の起源をトバ事変に関連づける向きもある。ヒトに寄生するヒトジラミ(人虱、Pediculus humanus)は2つの亜種、主に毛髪に寄宿するアタマジラミ(頭虱、P. h. capitis)と、主に衣服に寄宿するコロモジラミ(衣虱、P. h. humanus, P. humanuscorporis)に分けられる。近年の遺伝子の研究からこの2亜種が分化したのは約7万年前であることが分かっている。つまり、約7万年前にヒトが衣服を着るようになり、新しい寄宿環境に応じてコロモジラミが分化したと解釈される。そこで研究者らは、時期的に一致することから、トバ火山の噴火とその後の寒冷化した気候を生き抜くためにヒトが衣服を着るようになったのではないかと推定している。
近年では、ヨーロッパに進出したホモ・サピエンスはネアンデルタール人と、メラネシア方面へ進出したホモ・サピエンスはデニソワ人と交雑したという研究結果も発表されている。
なお、ヨーロッパ人と日本人の共通祖先の分岐年代は、7万年前±1万3000年であると推定されている。
印象的な遺伝的特徴(例えば皮膚の色)は、主に小集団が新たな環境へ移住した結果として起きた。これらの適応形質はホモ・サピエンスのゲノムの非常にわずかな部分によって引き起こされるが、皮膚の色のほかに鼻の形態や高高度地域で効率的に呼吸する能力など様々な形質を含む。
| 種類 | 分類 | 脳容積 (mL) |
|---|---|---|
| オランウータン | ヒト科 | 411 |
| ゴリラ | ヒト亜科 | 約500 |
| チンパンジー | ヒト族 | 394 |
| アウストラロピテクス・アフリカヌス | ヒト亜族 | 441 |
| ホモ・ハビリス | ヒト属 | 640 |
| ホモ・エルガステル | ヒト属 | 700-1100 |
| ホモ・エレクトゥス | ヒト属 | 1040 |
| ホモ・ハイデルベルゲンシス | ヒト属 | 1100-1400 |
| ホモ・ネアンデルターレンシス | ヒト属 | 1450 |
| ホモ・サピエンス・サピエンス | ヒト属 | 1350 |

心と行動の進化
人類の心と行動を進化させた要因については異なるいくつかの説がある。かつては脳の巨大化が二足歩行といった「知的な」行動の原因となったと考えられていた。しかし進化は目的論的には働かないと言う認識が深まりこの説は放棄された。何故ならヒトの祖先であるアウストラロピテクスはチンパンジー並みの440mlという非常に原始的な形態を示す脳を持つと同時に、完全に直立した下肢を持ち、大頭骨孔も頭の真下に位置し、二足歩行をしていた。脳の発達が人類進化の原点であるという20世紀初頭の考えは、アウストラロピテクスの発見により完全に否定されたのである。
知能の発達に関する説の一つはレイモンド・ダートの狩猟仮説である。動物を追い、効率よく狩りをするために予測や想像といった知性の発達が必要である。肉食による摂取エネルギーの増加は脳の増大を許容したかもしれない。狩猟仮説は戦争や暴力も狩猟活動の名残ではないかと予測する。しかし多くの生物で攻撃行動は捕食行動とは異なる部位の脳を活性化させる。また種内と種間の攻撃性は区別する必要がある。
一方、ドナ・ハートとロバート・サスマンは『ヒトは食べられて進化した』でヒトは長い間、捕食者ではなくてむしろ被食者であり、捕食を回避することが知能発達の選択圧になったと主張している。人類学者パスカル・ボイヤーは暗闇に対する恐怖、幽霊の錯覚のような認知的錯誤の一部が捕食者回避によって発達したのではないかと考えている。
米国・ユタ大学のデニス・ブランブル (Dennis Bramble) とハーバード大学のダニエル・リーバーマンは2004年、初期人類は、動物遺体から屍肉を集め、石を使って骨を割り、栄養価の高い骨髄を得ることを生息手段とする、一種の腐肉食動物であったとの仮説を提唱した。 人類は競合者に先駆けて動物遺体を手に入れるため、発汗による高い体温調整能力を始めとし、弾性のあるアキレス腱や頑丈な脚関節といった「速いピッチでの長距離移動の能力」を進化させ、広い地域を精力的に探し回る者として特化したとするものである。 このような適応の傾向と栄養価の高い食物が大きな脳の発達を可能にしたのではないかと説いた。
心理学者ニコラス・ハンフリーは複雑化する社会活動が重要な選択圧だと考えて社会脳仮説を提唱した。協力行動や騙し、騙しの発見などを行うには相手の心を読み、複雑な人間関係を理解する必要がある。心の理論の発達はこの一部であったかもしれない。霊長類学者ロビン・ダンバーは霊長類の大脳新皮質の大きさと様々な生活上の変数(食性、配偶システムなど)を比較し、群れの大きさとのみ相関があると指摘した。群れの巨大化は個人関係の複雑さに繋がる。社会脳仮説の支持者はダンバーの発見を証拠の一つと考えている。
認知考古学者スティーブン・ミズンは心のモジュール説を受け入れ、異なる神経構造を基盤に持ついくつかのモジュール化された心的機能(例えば言語能力、心の理論、直観的な物理の理解など)が個別に発達し、一般的知能が異なるモジュールの相互作用で完成したのではないかと考えている。
言語の利用
発声を扱ううえで、初期のヒト属(250-80万年前)の言語を扱う能力に関して著名な説がある。解剖学的に、350万年前ごろのアウストラロピテクスにおいて発達した二足歩行という特質が頭蓋骨に変化をもたらし、声道をよりL字形にしたと信じている学者もいる。頸部の比較的下の方に位置する声道や喉頭といった構造はヒトが作り出す多くの音声、特に母音を作るうえで必須な必要条件である。喉頭の位置に基づいて、ネアンデルタール人ですら現生人類が作り出す全ての音を完全に出すのに必要な解剖学的構造を具えていないと信じている学者もいる。さらに別の考え方では、喉頭の位置の低さは発声能力の発展とは無関係とされる。詳細は「言語の起源」を参照のこと。
道具の使用
道具の使用は知性の存在の象徴と解釈され、また道具の使用は人類の進化の特定の面(特に脳の継続的な増大)を刺激したかも知れないと推測されている。研究者は何百万年も続くこの負担の大きな器官の増大をまだ説明できていない。現代人の脳は20ワット(一日400キロカロリー)を消費し、人体の全消費量の20%にも達する。さらなる道具の使用は狩りと、植物よりエネルギーが豊富な肉の消費を可能にした。研究者は初期のヒト科が道具の作成と使用能力の増大を促すような選択圧のもとに置かれたと主張している。
初期の人類が道具を使い始めた正確な時期を特定するのは難しい。というのも原始的な道具(例えば鋭利な石)は人工物なのか自然にあるものか判別できないからである。アウストラロピテクスが400万年前に骨を道具として用いていた可能性を示す証拠があるが、これは議論の的である。
石器
石器は、約260万年前に初めてその証拠が現れる。東アフリカのホモ・ハビリスはいわゆる礫器、単純に打ち付けて割った丸い小石を用いていた。これは旧石器時代の始まりを意味する。旧石器時代は最終氷期の終焉期(約1万年前)に終わる。旧石器時代は前期(約35万年~約30万年前まで)、中期(約5万年~約3万年前まで)、後期に分けられる。約70万年から約30万年前の時代はアシュール文化 (cf. en:Acheulean) としても知られている。ホモ・エルガステル(またはホモ・エレクトゥス)は火打石と珪岩から大きな石斧を使っていた。最初(初期アシュール時代)には全く粗雑な作りであるが、のちには破片の縁で微妙に打ち付けることでより「加工された」道具を作った。
約35万年前にはより洗練されたルヴァロワ技法による石器作りが行われた。ルヴァロワ技法による石器の作成は完成予定の石器の形を正確に思い描かなくてはならず、抽象思考の証拠と考えられている。打ち付ける技術が洗練されると、鏝(こて)、スライサー、針なども作られるようになった。約5万年前にはネアンデルタール人と移住してきたクロマニョン人によって、より洗練され、特化された火打石やナイフ、刃物、毛皮などを剥くスキマーなどが作られた。この時期には骨からも道具が作られた。
火の利用
ヒトは、火を調理に使い、暖を取り、獣から身を守るのに使い、それによって個体数を増やしていった。火を使った調理は、ヒトがタンパク質や炭水化物を摂取するのを容易にした。火を利用することによって寒い夜間にも行動ができるようになり、あるいは寒冷地にも住めるようになり、ヒトを襲う獣から身を守れるようになった。
ヒト属による単発的な火の使用の開始は、約170万年から約20万年前までの広い範囲で説が唱えられている。最初期は、火を起こすことができず、野火などを利用していたものと見られるが、日常的に広範囲にわたって使われるようになったことを示す証拠が、約12万5000年前の遺跡から見つかっている。

ヒトの生活は、火とその明るさで大きな影響を受けた。夜間の活動も可能となり、獣や虫除けにもなった。また、当初は火を起こすのが難しかったため、火は集団生活で共用されるべきものとなり、それにより集団生活の必要性が増した。
火の使用は栄養価の向上にも繋がった。タンパク質は加熱することで、栄養を摂取しやすくなる。黒化した獣の骨から分かるように、肉も火の使用の初期から加熱調理されており、動物性タンパク質からの栄養摂取をより容易にした。加熱調理された肉の消化に必要なエネルギーは生肉の時よりも少なく、加熱調理はコラーゲンのゼラチン化を助け、炭水化物の結合を緩めて吸収しやすくする。また、病原となる寄生虫や細菌も減少する。
また、多くの植物には灰汁が含まれ、マメ科の植物や根菜にはトリプシンインヒビターやシアングリコーゲンなどの有毒成分が含まれる場合がある。また、アマ、キャッサバのような植物に有害な配糖体が含まれる場合もある。そのため、火を使用する前には植物の大部分が食用にならなかった。食用にされたのは種や花、果肉など単糖や炭水化物を含む部分のみだった。ハーバード大学のリチャード・ランガムは、植物食の加熱調理でデンプンの糖化が進み、ヒトの摂取カロリーが上がったことで、脳の拡大が誘発された可能性があると主張している。
実際、ホモ・エレクトゥスの歯や歯の付着物から、加熱調理無しには食べるのが難しい硬い肉や根菜などが見つかっている。
現代人と「偉大な飛躍」論争
約5万年から約4万年前にかけて、石器の使用は徐々に進歩していったと思われる。おのおのの段階(ホモ・ハビリス、ホモ・エルガステル、ホモ・ネアンデルターレンシスなど)は前の段階よりも高いレベルで始まり、後退したことはなかった。しかし一つの段階の中の技術の進歩は遅かった。言い換えると、これらの種は文化的に保守的であった。しかし、約5万年前以降、現生人類の文化は明らかに大きな速度で変わり始めた。『人間はどこまでチンパンジーか?』の著者ジャレド・ダイアモンドや他の人類学者はこれを「大躍進」と描写する。
現代の人間は丁寧に死者を埋葬し、隠れ家で衣類を作り、高度な狩猟技術をあみだし(穴を罠として使う、崖に動物を追い詰めるなど)、洞窟壁画を描き出した。この文化の変化のスピードアップは、現生人類、つまりホモ・サピエンスの誕生とその習性に関係しているようにみえる。集団の文化が進むと、異なる集団は既存の技術に新しい知識を取り入れる。釣り針、ボタンと骨製の針のような5万年以前は存在しなかった人工物は異なる人類の集団間の差異を示唆する。一般的にネアンデルターレンシスの集団は同時代の他のネアンデルターレンシス集団と同じような技術を用いていた。
理論的には現代の人間行動は次の4つの能力を含む:
- 抽象思考(具体的な例に依存しない概念)
- 計画(さらなるゴールを目指すためのステップを考える)
- 発想力(新たな解決法を見つける)
- 記号的な行動(儀式や偶像)
人類学者は現代的行動の具体例に以下を含める:
など。
しかしこれらの急激な出現が生物学的な革命的変化、「人間の意識のビッグバン」を意味するのか、より段階的な変化であったかの議論は続いている。コネチカット大学のサリー・マクブレアティとジョージ・ワシントン大学のアリソン・ブルックスは5万年以前の現代的行動の遺物を示し、革命説がアフリカの一部しかサンプルとしていないと主張して革命的進化はなかったと指摘した。
人類進化のモデル




今日、全ての人類はホモ・サピエンス・サピエンスに分類される。しかしこれはヒト属の最初の種ではない。ヒト属の最初の種、ハビリスは少なくとも200万年前に東アフリカで進化した。そして彼らは比較的短い時間でアフリカ各地に生息するようになった。ホモ・エレクトゥスは180万年以上前に進化し150万年前にはユーラシア大陸各地に広がった。実質的に全ての形質人類学者はホモ・サピエンスがホモ・エレクトゥスの子孫であることに同意する。人類学者はホモ・サピエンスが大陸各地で相互に関係しながら同時進行的にホモ・サピエンスになったのか(多地域進化説と呼ばれる)、東アフリカで現れた一派がユーラシア大陸各地のエレクトゥスと置き換わったのか(出アフリカ説、またはアフリカ単一起源説)で議論を行った。議論は未だ続いているが、大部分の人類学者は出アフリカ説を支持している。
多地域進化説
多地域進化説の支持者(主にミルフォード・ウォルポフと彼の同僚)は、ある程度の遺伝子流動があればヨーロッパとアジアの異なる地域で並行的に現生人類の進化が可能であったろうと主張した。古代ヨーロッパと中国のホモ・サピエンスの形態的な類似性と、それぞれの地域の古代と現代のホモ・サピエンスの類似性は地域的な進化を支持しているとウォルポフは主張する。彼らはさらにこの説が表現型多型のクラインパターンと一致しているとも主張する。
出アフリカ説
クリス・ストリンガーとピーター・アンドリューズによって発展した出アフリカ説によれば、分子系統解析の進展(いわゆるミトコンドリア・イブやY染色体アダムなど)によって、現代のホモ・サピエンスは14 - 20万年前に共通の祖先を持つことがわかり、ホモ・サピエンスは7万から5万年前にアフリカから外へ移住し始め、結局ヨーロッパとアジアで既存のヒト属と置き換わった。出アフリカ説はミトコンドリアDNAを用いた最近の研究によっても支持された。133種類のミトコンドリアDNAを用いた系統樹の分析の結果、彼らは人類が(のちにミトコンドリア・イブと呼ばれる)アフリカ女性の子孫であると結論した。ただしミトコンドリア・イブは全人類の「ミトコンドリアDNAの」祖先であり、人類がただ一人の女性あるいは夫婦のみに由来するという意味でも、この女性が最初のホモ・サピエンスという意味でもない。
ミトコンドリアDNAの分析では、現代人の共通祖先の分岐年代は14万3000年前±1万8000年であり、ヨーロッパ人と日本人の共通祖先の分岐年代は、7万年前±1万3000年であると推定された。
出アフリカの回数が一度であったか、複数回であったかには議論があるが、ユーラシアとオセアニアの住民はみな共通したミトコンドリアDNAの系統に属していることは、複数回出アフリカ説に対する重要な反証である。ほとんどの研究は、一度だけの出アフリカが、アフリカ以外の全人類の起源となった可能性を示唆する。
出アフリカのルートも複数説あるが、近年、遺伝学的、言語学的、考古学的な証拠の支持を得ているのは、およそ7万年前にアフリカ東部の突端であるいわゆるアフリカの角からアラビア半島に渡った、「南方出アフリカ説」である。
二つのモデルの比較
二つのモデルは非常に異なる。リチャード・リーキーは次のようにこの違いをまとめている。
- 多地域進化モデルは集団の置換が起きず、移住もわずかで、旧世界各地でホモ・サピエンスの進化的傾向があったと述べている。一方、出アフリカモデルでは一カ所でのホモ・サピエンスが進化し、そして旧世界全域への広範な人口移動と既存の前現代的な集団との置換が起きると述べる。
多地域モデルは化石記録が現在見えるような地域ごとの解剖学的特徴を示さなければならないと提案する。そして人種的な違いは根深く、200万年遡ると主張する。アフリカ単一モデルでは化石記録は時間に従った連続性を示すとは主張しない。そうではなくて、以前の地域ごとの化石記録の特徴は現代アフリカ人的な特徴を持つ化石史料によって置き換えられる。人種的な違いは浅く、比較的短い期間で人種的差異は進化したと主張する。
アクア説(水生類人猿説)
現在のところ主流派からはあまり支持されていないが一部でこのアクア説(水生類人猿説)が唱えられている。 ヒトがチンパンジー等の類人猿と共通の祖先から進化する過程で、水生生活に一時期適応することによって直立歩行、薄い体毛、厚い皮下脂肪、意識的に呼吸をコントロールする能力といった他の霊長類には見られない特徴を獲得したとするもので、解剖学者と海洋生物学者が提唱し、脚本家であるエレイン・モーガンの著作で知られるようになった仮説である。
研究者
ここでは、人類進化の研究分野において特筆性の高い人物を列記する。
- チャールズ・ダーウィン - 1809年生まれ。イングランド人。博物学者。進化的変化を通して種が誕生することの重要な証拠を提示した。
- ロバート・ブルーム - 1866年生まれ。スコットランド人。医者、古生物学者。オレンジ自由国(現・南アフリカ共和国の一角)のスタルクフォンテインで「ミセス・プレス」の発見を導いた。
- レイモンド・ダート - 1893年生まれ。オーストラリア人。解剖学者、古人類学者。オレンジ自由国のタウングでアウストラロピテクスを発見した。
- アリスター・ハーディ - 1896年生まれ。イギリス人。海洋生物学者。水棲人類仮説を提唱した初期の一人。
- ルイス・リーキー - 1903年生まれ。イギリス人。アフリカで活動した古生物学者で、人類学者、考古学者。アフリカにおける人類の進化的発達を立証するのに重要な役割を果たした。
- メアリー・リーキー - 1913年生まれ。イギリス人。古生物学者。アフリカで発掘を行い、ラエトリ遺跡などを発見した。
- ルイジ・ルーカ・カヴァッリ=スフォルツァ - 1922年生まれ。イタリア人。集団遺伝学者、言語・生物学者。ヒトの拡散を生物学的側面だけではなく、文明・文化的側面からもアプローチし、その整合性を指摘した。
- フィリップ・V・トビアス - 1925年生まれ。南アフリカ連邦(南アフリカ共和国の前身)の人。古人類学者。人類の進化研究の世界的権威の一人。
- ミルフォード・H・ウォルポフ - 1942年生まれ。アメリカ人。古人類学者。多地域進化説の主要な支持者。
- ヘンリー・マクヘンリー - 1944年生まれ。アメリカ人。人類の進化、特に二足歩行の起源を専門とするアメリカの人類学者、古人類学者。
- クリス・ストリンガー(クリストファー・ブライアン・ストリンガー;Christopher Brian Stringer) - 1947年生まれ。イギリス人。物理人類学者。アフリカ単一起源説の主要な支持者。
- ジェフリー・H・シュワルツ - 1948年3月6日生まれ。アメリカ人。形質人類学者、生物人類学者。
- エリク・トリンカウス - 1948年12月24日生まれ。アメリカ人。古人類学者。専門はネアンデルタール人とホモ・サピエンスの進化。
- スバンテ・ペーボ - 1955年生まれ。スウェーデン人。進化遺伝学者。ネアンデルタール人のDNAの分析などを行っている。
- アラン・テンプルトン - 生年不詳(20世紀後期前半ごろ)。アメリカ人。遺伝学者、統計学者。多地域進化説の支持者。
- スペンサー・ウェルズ - 1969年生まれ。アメリカ人。分子生物学者、人類遺伝学者。全世界のあらゆる地域、あらゆる民族の遺伝子データを集め、人類の進化と拡散の過程を明らかにしていくことを目標としたジェノグラフィック・プロジェクトの総指揮者。
種リスト
ヒト属に属する種は「ヒト属:分類」を参照のこと。
脚注
注釈
- Googleマップ
参考文献
- 池田清彦『38億年 生物進化の旅』新潮社、2010年2月25日。OCLC 534633602。ISBN 4-10-423106-1、ISBN 978-4-10-423106-5。
- 長谷川政美『系統樹をさかのぼって見えてくる進化の歴史─僕たちの祖先を探す15億年の旅』ベレ出版〈BERET SCIENCE〉、2014年10月24日。OCLC 897123465。ISBN 4-86064-410-7、ISBN 978-4-86064-410-9。
この節で示されている出典について、該当する記述が具体的にその文献の何ページあるいはどの章節にあるのか、特定が求められています。
|
- Flinn, M. V., Geary, D. C., & Ward, C. V. (2005). Ecological dominance, social competition, and coalitionary arms races: Why humans evolved extraordinary intelligence. Evolution and Human Behavior, 26, 10-46. Full text. (PDF, 345 KiB)
- Rozsa L 2008. The rise of non-adaptive intelligence in humans under pathogen pressure. Medical Hypotheses, 70, 685-690.
- Wolfgang Enard et al. (2002-08-22). “Molecular evolution of FOXP2, a gene involved in speech and language”. Nature 418: 870.
- DNA Shows Neandertals Were Not Our Ancestors
- J. W. IJdo, A. Baldini, D. C. Ward, S. T. Reeders, R. A. Wells (October 1991). “Origin of human chromosome 2: An ancestral telomere-telomere fusion”. Genetics 88: 9051–9055.—two ancestral ape chromosomes fused to give rise to human chromosome 2.
- Ovchinnikov, et al. (2000). “Molecular analysis of Neanderthal DNA from the Northern Caucasus”. Nature 404: 490. doi:10.1038/35006625.
- Heizmann, Elmar P J, Begun, David R (2001). “The oldest Eurasian hominoid”. Journal of Human Evolution 41 (5): 463. doi:10.1006/jhev.2001.0495.
- Haldane, John Burdon Sanderson (23 July 1955). “Origin of Man”. Nature (Nature Publishing Group (NPG)) 176 (169). doi:10.1038/176169a0.
- BBC: Finds test human origins theory. 2007-08-08 Homo habilis and Homo erectus are sister species that overlapped in time.
関連項目
|
|
|