
染色体の定義
歴史的背景から、染色体という語には複数の定義がある。
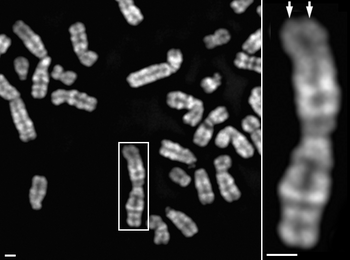
- 原義では、細胞分裂期に観察される棒状の構造体を指す。染色体の形態として一般的に認識されている構造は、この分裂期のものである(図1)。
- 広義では、形態や細胞周期に関わらず、真核細胞にあるゲノムDNAとタンパク質の巨大な複合体を指す場合がある。
- さらに広義には、細菌や古細菌あるいはミトコンドリアなどの細胞小器官が持つゲノムを含めて染色体と呼ぶこともある(核様体の項参照)。ウイルスのゲノムも染色体と呼ぶ場合がある。
染色体の構造

染色体の最も基本な構成要素は、DNAとヒストンである。分裂期の染色体は一対の姉妹染色分体から構成され、それぞれの染色分体には長いDNA一分子が含まれている。DNAは酸性であり、塩基性タンパク質のヒストンとの親和性が高い。DNAとヒストンの重量比は、ほぼ1:1である。
染色体の最も基本的な構造はヌクレオソームである(図2)。4種のコアヒストン(H2A, H2B, H3, H4)が2つずつ集まってヒストン8量体を形成し、146塩基対の2重鎖DNAを左巻きに巻きつける(DNA超らせんの項参照)。ヌクレオソームとヌクレオソームをつなぐDNAはリンカーDNAと呼ばれ、そこにはリンカーヒストン(ヒストンH1など)が結合する。ヌクレオソーム構造はさらに凝集して直径30 nmの繊維を形成すると考えられているが、その構造についてはいまだ定説がない。分裂期にはいると、光学顕微鏡下で観察可能な棒状の構造体(第一の定義における染色体)に変換される。この染色体凝縮過程には、コンデンシン複合体やトポイソメラーゼ II が関与する。
クロマチンには、大きく分類してユークロマチン (euchromatin) とヘテロクロマチン (heterochromatin) の二種類がある。ユークロマチンはクロマチン構造がゆるまっており、転写されている遺伝子はこの部分に多く存在する。ヘテロクロマチンは密に凝集しており、この領域ではあまり転写が起きていない。ヘテロクロマチンは更に次の二つに分類することができる。遺伝子の発現はほとんど見られない構成的ヘテロクロマチン (constitutive heterochromatin) と、条件によっては遺伝子の発現が見られる条件的ヘテロクロマチン (facultative heterochromatin) がある。前者は主にセントロメア付近にあり、この領域の DNA は繰り返し配列に富む。
染色体の各部位の呼称

分裂期の染色体は一対の姉妹染色分体からなる(図3)。染色分体どうしがより強固に接着している領域はセントロメアと呼ばれる。分裂期にはセントロメア上に形成されるキネトコアに微小管が結合し、染色分体を両極へ牽引する。セントロメアをはさんで長い側を長腕、短い側を短腕という。染色体の末端部はテロメアと呼ばれる特有の構造をしている。
染色体を構成するタンパク質因子
染色体にはヒストンの他にも多くのタンパク質因子が結合している。RNAポリメラーゼのような基本転写因子と呼ばれるタンパク質複合体や、特定の遺伝子座に結合しその遺伝子の発現を制御するもの、クロマチンの状態を維持または変化させるものなどがある。また、トポイソメラーゼと呼ばれる一群の酵素は、DNA超らせん状態を制御する。染色体の高次構造を制御する因子の中で代表的なものには、染色体凝縮に関わるコンデンシンや姉妹染色分体の接着に関与するコヒーシンがある。
細胞周期における染色体の挙動

有糸分裂の最初のステージ(前期)では、核膜を保持したまま、クロマチンが凝縮を開始する。前中期では、核膜が崩壊し、染色体凝縮がさらに進行する。凝縮の最も進んだ分裂中期では、二つの染色分体(姉妹染色分体)がセントロメアでより強固に結合した形態をとる。細胞の両極から伸びた長い微小管(紡錘糸)はキネトコアに結合する(図4)。分裂後期にはいると、姉妹染色分体間の接着が解除され、紡錘糸は各染色分体を細胞の両極に向けて引き離す。(染色体分離)。こうして、最終的に各娘細胞は1セットの染色分体を受け継ぐ。細胞分裂が完了すると、染色分体は再び脱凝縮して細胞核内に収納される。
核型

ある生物の染色体を調べたいとき、コルヒチン等の薬剤で細胞を処理し細胞分裂をM期で停止させてからギムザ等の染色を施し、凝縮した染色体の数と形状を観察する。こうして撮影された染色体を並べたものが、核型(karyotype カリオタイプ、karyogram カリオグラム とも呼ばれる)である。これを調べて分類学的検討などを行うことを核型分析という。
カリオタイプは種ごとに一定である。例えば、ヒトの二倍体細胞は、22対の常染色体と1対の性染色体、計46本の染色体を持つ(図5)。性染色体の組み合わせは女性では2本のX染色体、男性ではX染色体とY染色体1本ずつとなっている。女性の2本のX染色体のうちの片方は不活性化されており、顕微鏡下ではバー小体として観察される。
有性生殖を行う多くの種は、二倍体 (2n) の体細胞 と一倍体 (1n) の配偶子を持つ。雄由来の配偶子と雌由来の配偶子が接合(受精)すると、二倍体の接合子(受精卵)となり、体細胞分裂を繰り返して個体をつくりあげる。すなわち、二倍体の体細胞が有する2セットの相同染色体のうち、1セットは父親から、もう1セットは母親から由来する。一倍体の配偶子をつくるための特殊な細胞分裂は、減数分裂と呼ばれる。減数分裂の過程では母親と父親に由来する相同染色体は交叉を起こして遺伝情報を交換する。このように、片親からの染色体をそのまま次の世代に渡すのではなく、世代を経るたびに常に新しい遺伝情報の組み合わせが作られるようになっている。無性生殖で増殖する種の多くは染色体を1セットしか持たない。
なお、男性の持つY染色体はかつて、その大きさや遺伝子の位置などがX染色体と異なることから、減数分裂時の遺伝子の組み換えを起こさない、変異しづらい不活性なものとされてきた。しかし最近では、Y染色体においてもX染色体との交叉による乗り換えが起こっていると考えられている。またY染色体内で、自身の遺伝子の位置が入れ替わっていることが明らかになるなど、実際にはY染色体の変異は比較的頻繁に起きていることがわかっている。
生物種による染色体数の違い
|
|
染色体研究の歴史
- 1842年、ネーゲリ(Carl Nageli)が、染色体を発見。
- 1865年、メンデル(Gregor Mendel)がメンデルの法則を発表。
- 1869年、ミーシャー(Friedrich Miescher)が、ヌクレイン(今日のDNA)を発見。
- 1882年、フレミング (Walther Flemming)が有糸分裂の詳細を記載。
- 1888年、ヴァルデヤー ( H. W. G. von Waldeyer-Hartz)が「染色体(chromosome)」を命名。
- 1900年、 ド・フリース(Hugo de Vries)、チェルマク(Erich von Tschermak)、コレンス(Carl Correns)によるメンデルの法則の再発見。
- 1902年、サットン(Walter Sutton)による染色体説の提唱。
- 1920年代、モーガン(Thomas Hunt Morgan)らによる染色体説の実証(下記参照)。
- 1944年、アベリー(Oswald Avery)らによる肺炎双球菌の形質転換実験(アベリー-マクロード-マッカーティの実験)。
- 1952年、ハーシー(Alfred Hershey)らによるブレンダー実験(ハーシーとチェイスの実験)。
- 1953年、ワトソン(James Watson)、クリック(Francis Crick)によるDNA二重らせんモデルの提唱。
- 1956年、アーサー・コーンバーグ(Arthur Kornberg)によるDNAポリメラーゼの発見。
- 1974年、オリンズ(A. Olins & D. Olins)、コーンバーグ(Roger Kornberg)らによるヌクレオソームの発見。
- 1978年、ブラックバーン(Elizabeth Blackburn)らによるテロメア配列の同定。
- 1980年、カーボン(John Carbon)らによる機能的セントロメア配列の同定。
- 1996年、アリス(Charles David Allis)らによるヒストンアセチル化酵素の同定。シュライバー(Stuart Schreiber)らによるヒストン脱アセチル化酵素の同定。
- 1997年、リッチモンド(T. J. Richmond)らによるヌクレオソームの高解像度結晶構造解析。
- 2000年、ヒトゲノムドラフト配列の発表。
ハエ目昆虫のショウジョウバエやユスリカの幼虫のだ腺染色体(唾液腺細胞中の染色体)は通常の体細胞の染色体とは異なり、多糸染色体とよばれている。この染色体は例外的に、顕微鏡下でよく観察することができる。モーガンらによる初期の遺伝子研究では、主にショウジョウバエのだ腺染色体を材料として染色体上の遺伝子の位置が決定され、染色体地図が作成された。これらの成果は近年のホメオボックス遺伝子などショウジョウバエを材料とした遺伝子研究の基礎をなすものとなったばかりでなく、遺伝学全般の基礎をなしていると言える。
脚注
注釈
参考図書
- B. Alberts他 著(中村桂子・松原謙一 監訳)『細胞の分子生物学 第6版』ニュートンプレス、2017年。
- B. Alberts他 著(中村桂子・松原謙一 監訳)『Essential 細胞生物学 第4版』南江堂、2016年。
- D. Morgan 著(中山敬一・啓子 翻訳)『カラー図説 細胞周期』メディカルサイエンスインターナショナル、2008年。
- A.T. Sumner 著(福井希一・真庭理香 翻訳)『クロモソーム:構造と機能』大阪公立大学共同出版会、2007年。
- 平岡泰・原口徳子 編『染色体と細胞核のダイナミクス』化学同人、2013年。
- 平野達也・胡桃坂仁志 編『実験医学増刊号「教科書を書き換えろ!染色体の新常識」』羊土社、2018年。
資料
ヒトの染色体の情報
| 染色体
番号 |
遺伝子数
(個) |
塩基対数
(bp) |
|---|---|---|
| 1 | 2,610 | 279,000,000 |
| 2番染色体 (ヒト) | 1,748 | 251,000,000 |
| 3番染色体 (ヒト) | 1,381 | 221,000,000 |
| 4番染色体 (ヒト) | 1,024 | 197,000,000 |
| 5番染色体 (ヒト) | 1,190 | 198,000,000 |
| 6番染色体 (ヒト) | 1,394 | 176,000,000 |
| 7番染色体 (ヒト) | 1,378 | 163,000,000 |
| 8番染色体 (ヒト) | 927 | 148,000,000 |
| 9番染色体 (ヒト) | 1,076 | 140,000,000 |
| 10番染色体 (ヒト) | 983 | 143,000,000 |
| 11番染色体 (ヒト) | 1,692 | 148,000,000 |
| 12番染色体 (ヒト) | 1,268 | 142,000,000 |
| 13番染色体 (ヒト) | 496 | 118,000,000 |
| 14番染色体 (ヒト) | 1,173 | 107,000,000 |
| 15番染色体 (ヒト) | 906 | 100,000,000 |
| 16番染色体 (ヒト) | 1,032 | 104,000,000 |
| 17番染色体 (ヒト) | 1,394 | 88,000,000 |
| 18番染色体 (ヒト) | 400 | 86,000,000 |
| 19番染色体 (ヒト) | 1,592 | 72,000,000 |
| 20番染色体 (ヒト) | 710 | 66,000,000 |
| 21 | 337 | 45,000,000 |
| 22 | 701 | 48,000,000 |
| X | 1,098 | 163,000,000 |
| Y | 78 | 51,000,000 |
- 桁数を揃える為に、首位が「0」であっても表示してある。
- 月刊科学雑誌Newton2006年2月号『「性」を決めるカラクリ XY染色体』による。(複数の情報を合わせたものであり、中立的なため)
- 資料によって値が大きく変わる事もある。(特に、X・Y染色体の遺伝子数)
- 日本語の資料はこちら。(京都大学大学院 生命科学研究科 生命文化学研究室による。上の表とは異なる部分もある。)
- 最新の情報はNCBIのサイト(英語)で公開されている。