
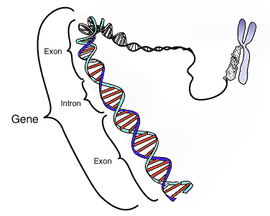
概念
分子生物学における最狭義の遺伝子はタンパク質の一次構造に対応する転写産物 (mRNA) の情報を含む核酸分子上の特定の領域=構造遺伝子(シストロン)をさす。転写因子結合部位として、転写産物の転写時期と生産量を制御するプロモーターやエンハンサーなどの隣接した転写調節領域を遺伝子に含める場合もある(→オペロン)。ちなみに、語感が似る調節遺伝子とは上記の転写因子のタンパク質をコードしたれっきとした構造遺伝子である。 しかし、転写産物そのものが機能を持ち、タンパク質に翻訳されない、転移RNA (tRNA) やリボソームRNA (rRNA) 、機能性ノンコーディングRNAに対応する遺伝情報が、タンパク質構造遺伝子と同程度の数をもつことが報告され、狭義の遺伝子に含められるようになっている。
古典的な遺伝子の定義は、ゲノムもしくは染色体の特定の位置に占める遺伝の単位(→遺伝子座)であり、構造は変化しないと考えられていた。しかし突然変異やトランスポゾン(可動性遺伝子)の発見、抗体産生細胞で多種の抗体を作り出すための遺伝子再編成の発見などから、分子生物学的実験対象としての遺伝子の概念はたびたび修正を余儀なくされた。
また同じ生物学内でも進化論や集団遺伝学、進化ゲーム理論での議論で用いられる遺伝子という単語は、上記の構造遺伝子やDNA上の領域あるいは遺伝子座とは相当に異なる概念を内包しており、混同してはならない(例:リチャード・ドーキンスの著書表題『The Selfish Gene(利己的な遺伝子)』)。こちらは、自然選択あるいは遺伝的浮動の対象として集団中で世代をまたいで頻度を変化させうる情報単位である。メンデル遺伝的な面をもつもののほか、表現型に算術平均的影響を与える量的形質遺伝子、遺伝情報の突然変異や組み換えに対応する無限対立遺伝子モデルなど、理論的でありながら、即物的な分子生物学の側面を包含した考え方である。これを模倣し、文化進化の文脈で用いられるミームは集団遺伝学における遺伝子のアナロジーである。
遺伝子という言葉は、「遺伝する因子」としての本来の意味を超えて遺伝子産物の機能までを含んで用いられる場合があり、混乱を誘発している。後者の典型例としては、遺伝しない遺伝子を使った遺伝子治療などがあげられる。さらに遺伝子やDNAという言葉は、科学的・神秘的といったイメージが先行し、一般社会において生物学的定義から離れた用いられ方がされていることが多い。それらの大半は通俗的な遺伝観を言い換えたものに過ぎない。一般雑誌などでは疑似科学的な用法もしばしば見受けられる。
機能

遺伝子はDNAが複製されることによって次世代へと受け継がれる。複製はDNAの二重らせんが解かれて、それぞれの分子鎖に相補的な鎖が新生されることで行われる。
本質的には情報でしかない遺伝子が機能するためには発現される必要がある。発現は、一般に転写と翻訳の過程を経て、遺伝情報(= DNAの塩基配列)がタンパク質などに変換される過程である。こうしてできたタンパク質が、ある場合は直接特定の生体内化学反応に寄与して化学平衡などに変化をもたらすようになり、ある場合は他の遺伝子の発現に影響を与え、その結果形質が表現型として現われてくる。転写はDNAからRNA(mRNAやrRNAなど)に情報が写し取られる現象であり、翻訳はmRNAの情報を基にタンパク質が合成される過程である。この過程はセントラルドグマとも呼ばれる。
遺伝子の発現
遺伝子発現に関する多くの知見は真核生物ではなく細菌である大腸菌をモデル生物とした実験から得られてきた。
真核生物の遺伝子の一般的な働き方
核内では様々なDNA結合特異性を持った転写調節因子の転写調節領域への結合や、DNAのメチル化状態などで遺伝子の活性が制御されている。DNAからRNAポリメラーゼによってRNAへと転写された転写産物はmRNA前駆体と呼ばれる。
これが、5'末端へのキャップ構造の付加やスプライシング、3'末端の切断、ポリA鎖の付加といった作用を受けてmRNAとなる。mRNAは転写の場である核から核膜孔を通過し細胞質へ運ばれる。
細胞質では、キャップ構造を認識する蛋白質や翻訳開始因子との作用によりリボソームがmRNAに結合する。リボソーム上では、コドンに対応したアンチコドンを持ったアミノアシルtRNAがAサイトに結合することで塩基配列からアミノ酸配列への遺伝情報の翻訳が行われる。Pサイトに結合しているペプチジルtRNAから、アミノ酸が連なったポリペプチドがAサイトのtRNAに付加され、これがPサイトに移動することが繰り返される。
翻訳されたアミノ酸配列はその一次構造に依存した立体構造をもつ蛋白質へと折り畳まれる。蛋白質の機能はその立体構造によって規定されており、正常な構造をもつ蛋白質がさらに糖鎖の付加やリン酸化といった翻訳後の修飾をうけて最終的な遺伝子産物となることもある。
遺伝子研究
一般に、遺伝子研究とは遺伝学、分子生物学、ゲノミクスなどの研究を指す。集団遺伝学や進化遺伝学は含めないことが多い。
遺伝子研究はメンデル・モーガンの古典遺伝学に始まった。古典遺伝学における遺伝子研究はメンデルの行ったような交雑実験と表現型の観察を中心とし、遺伝子は遺伝情報を担う粒子の概念として扱われた。
分子生物学黎明期では主に大腸菌やファージを用いて、DNAを直接扱う形質転換実験や、DNA塩基配列からの遺伝子発現機構の解析などが行われた。現在では様々なモデル生物に研究対象が拡大している。これは、遺伝子の実体がほとんど全生物において『DNAである』ことによる(DNAを扱えればいかなる生物でも分子生物学的実験は行える)。
突然変異の表現型から遺伝子機能を推定する正の遺伝学はマウスなどでは行いづらく、先に遺伝子を同定してから変異体を作成する逆遺伝学という手法が生まれた。逆遺伝学の先にゲノムプロジェクトがあり、さまざまな生物種で進行または終了している。ゲノムプロジェクトによって遺伝子の数を有限に規定することができる。
DNAの構造決定とゲノムプロジェクトは遺伝子研究にパラダイムシフトをもたらした。シーケンシング(塩基配列の解析)技術は2009年現在も飛躍的に進歩し続けており、高速かつ低コストにゲノム全体を網羅的に解析できるようになりつつある。単に塩基配列を知るというレベルではなく、個体差を比較したり、遺伝子の発現パターンをプロファイリングしたりといった、従来は困難と認識されてきた研究も現実的に可能となってきた。in vivoにおける遺伝子の機能、すなわち『遺伝子はどのように生物体で機能しているのか』という問いへの答えが明らかになりつつある。
遺伝子操作の概要
遺伝子の機能を調べるにはいくつかのテクニックが必要である。生物体内 (in vivo) における特定の遺伝子はいくつかのコピーが存在するものの、その遺伝子が何を意味しているのか、発現するとどうなるのか、変異が起こればどうなるのかを調べることは困難である。したがって、その遺伝子のみを取り出して、遺伝子の特性を生物体外 (in vitro) で調べる必要がある。それらの過程には
の三段階を経る。クローニングや発現の前には、サブクローニングや発現ベクターへの遺伝子の導入といったプロセスを経ることもある。
クローニング
クローニングとは、遺伝子のクローンを作成する実験である。遺伝子のクローンを作成するにはある程度の配列がわかっていることを前提に現在2つの方法が実用化されている。
- ゲノムDNAを制限酵素で切断し、サザンブロッティングで目的遺伝子を含む配列を同定する方法
- PCR法を用いて目的の遺伝子配列を増幅する方法
遺伝子配列がわからない場合には、目的のタンパク質を対象生物内から精製し、タンパク質N末端配列を決定した後、ミックスプライマー(アミノ酸とその遺伝暗号に対応するパターン全てを含む複数種のプライマー、詳しくは当該記事にて)を用いてクローニングができる。
上記いずれのケースにおいても、単一のDNA配列のみを増幅した、あるいは精製したのみではヌクレアーゼによって分解を受けてしまう。したがって、目的DNA配列をクローニングベクターに導入し、大腸菌を用いてベクターを増幅することを含めてクローニングという実験が完結する。
細菌は遺伝子に介在配列を持たないためにDNAから遺伝子をクローニングすることが可能だが、真核生物の場合はイントロンをのぞいたエクソン部分のみを抽出する必要がある。これはスプライシング後のmRNAを精製し、逆転写PCR (RT-PCR) を行うことによってクローニングが可能となる。
シークエンシング
シークエンシングとはDNAの塩基配列の並びを決定する実験を意味する。シークエンシングを行うには、やはりある程度の配列が判明している必要があるが、クローニングが可能であれば特に問題はない。シークエンシングにはかつてマクサム - ギルバート法が用いられていたが、現在はサンガー - クルソン法(ジデオキシヌクレオチド鎖終結法)の変法である『ダイターミネーターサイクルシークエンシング法』が一般的である。
2009年現在もっとも一般的に使われているシークエンシング技術では、1つのプライマーから1,200塩基対の配列が一回の実験で決定可能である。しかし「次世代型シーケンサー」と総称される高速解読装置が複数のメーカーから発表されており、これらを用いれば従来よりはるかに大量の配列情報を短時間に得ることができる。たとえばアプライドバイオシステムズ社のSolidシステムでは一回の解析で30億塩基対の解読が可能という。
過剰発現
タンパク質を用いた実験を行うには、
- 生物内からタンパク質を精製する、
- 過剰発現系(かじょうはつげんけい)を用いて大腸菌等ベクターの宿主に目的タンパク質を大量に発現させる
という二通りの方法がある。生物内からタンパク質を精製するには、大量のサンプルが必要であり、タンパク質精製のテクニックが必要であるために一般的な技術とは言いがたい。一方過剰発現系を用いれば、誰でも簡単に目的のタンパク質を大量に得ることができる。
最も一般的な過剰発現系には、発現ベクター中の大腸菌のlacZプロモーター配列の下流に、クローニングした遺伝子を導入する方法がある。この方法では、IPTGという物質を用いてlacZプロモーター下流の遺伝子を大腸菌内で発現させることができる。転写されたmRNAはその後、大腸菌のリボソームで翻訳され、大量にタンパク質を生産する。
このようにして生産したタンパク質を用いて、酵素であれば活性を測定したり、DNA結合タンパクであればDNA結合実験を行ったりとタンパク質の実験が可能である。現在、ポストゲノムと言われる分野の主流はこの過剰発現系を用いたものである。しかしながら、大腸菌発現系では多くの問題を抱えており、現在大腸菌以外にも多くの発現宿主が開発されている(細菌:細胞外酵素作成型(Bacillus属を用いたもの)真核生物:出芽酵母、動物細胞、ヒト細胞などなど)。
以上のような方法と比較して、要するコスト及び時間を低下させることが可能な、コムギ胚芽を利用した無細胞タンパク質合成系という新たな方法も研究されている。
遺伝子研究の応用
遺伝子導入とは、上記にあげた遺伝子の実験系を用いて目的遺伝子の宿主でない他生物にクローニングしたベクターを導入し、その遺伝子が有効な形質を発現できるように仕向けることである。例えば、特定の除草剤に対して耐性を持つような作物や、霜害を防ぐ糖タンパクを生産できる作物(アイスマイナス)などはその一例である。
しかしながら、導入したベクターが花粉などを通じて拡散し、除草剤耐性を持っていなかった雑草にまでそうした形質が導入される危険性を指摘され、このような遺伝子組み換え実験は厳しく規制された状況である。遺伝子組み換え実験による物理的規制は
- P1:一般的な実験室
- P2:病原性を扱うような実験室
- P3:バイオクリーンルーム(室内に流入する空気は全てHEPAフィルタ(0.22μm方形のフィルター)を通している)
- P4:遺伝子組み換え対象生物に触れずに実験できる実験室
というランクが付けられており、危険な遺伝子組み換え実験を行う場合にはそれ相応の規制のランクの敷かれた実験室で行うよう義務付けられている。
なお、遺伝子組み換えの規制に関して、1975年、アメリカで開かれたアシロマ会議において基本的なガイドラインが決められた。遺伝子組み換えによって誕生した生物の取り扱いについては、2003年、生物の多様性に関する条約としてカルタヘナ議定書が締結された。
歴史
現在の遺伝子の概念はメンデルによって定義される。彼はエンドウのいくつかの表現型に注目した交雑実験を行い、表現形質が分離することを発見する(1865年 →メンデルの法則)。これを説明するために形質を伝える因子たる「遺伝粒子」を考え、これが現在の遺伝子の基となっている。それまで形質は液体のように混じりあっていくと考えられていた。しかし、長らくメンデルの法則は不評で、1900年に再発見されるまで理解されなかった。
細胞学者たちは減数分裂の様子を観察し、対になった染色体が一つずつになり、接合後に再び対を作るという染色体の挙動が、遺伝子のそれと同じであることを発見した(→染色体説)。ショウジョウバエの突然変異を用いた遺伝学的実験によりそれが明らかにされた。
染色体はタンパク質と核酸からできていることが明らかにされたが、当時はタンパク質が遺伝子の正体であると考えられていた。多数の種類があるタンパク質に比べ、核酸はあまりにも多様性が低いと考えられていたためである。実際、100bpのDNAの情報量は約10の60乗 (4100) であるのに対し、100個のアミノ酸で構成される蛋白質の情報量は10の130乗 (20100 = 2100 × 10100) と甚だ差が激しい。
しかし、肺炎双球菌やファージを用いた実験で DNA が遺伝子の正体であることが実証され、そのすぐ後に DNA の構造が解明された。DNA の二重らせん構造は遺伝子の性質と非常によく一致していた。
メンデルの法則発見から二重らせん構造発見までの歴史
- 1865年 グレゴール・ヨハン・メンデルがエンドウの交雑実験の結果を発表。(→メンデルの法則)
- 1869年 フリードリッヒ・ミーシェルが膿の細胞抽出液からDNAを発見する。
- 1900年 メンデルの投稿した論文がユーゴー・ド・フリース(オランダ)、カール・エーリヒ・コレンス(ドイツ)、エーリヒ・フォン・チェルマク(オーストリア)によって再発見される。この再発見者の一人フリースはパンゲン説を推し、細胞内で形質を伝達する物質をパンゲンと仮定する。
- 1903年 ウォルター・S・サットンがパンゲンが染色体上にあることを提唱した。(→染色体説)
- 1909年 ウィルヘルム・ヨハンセンはメンデルの指摘した因子をフリースの名づけたパンゲン (pangen) から『gene(遺伝子)』と呼ぶことを提案した。
- 1910年 トーマス・ハント・モーガンがショウジョウバエの交雑実験を始める。
- 1921年 DNAのテトラヌクレオチドモデルを解説した論文が発表される(J. Biol Chem. 48:119〜125)。当時、遺伝物質は多様性に富んだポリペプチド(タンパク質)であり、テトラヌクレオチドはその保護の役割を果たしていると考えられていた。
- 1922年 モーガンらのグループによってショウジョウバエの4つの染色体上に座している50個の遺伝子の相対位置が決定され、発表される。
- 1927年 ハーマン・J・マラーがX線は遺伝子に突然変異を導入することを指摘する。
- 1934年 カスパーソンがDNAは生体高分子であることを示し、テトラヌクレオチドモデルが誤りであることが証明される。
- 1935年 マックス・デルブリュックらは、遺伝子は物質的単位であることを提案した。
- 1941年 ビードルとタータムが『一遺伝子一酵素説』(1つの遺伝子は1つの酵素をコードしている)を発表。
- 1944年 フレデリック・グリフィスの肺炎双球菌の形質転換実験(グリフィスの実験)を元にした、オズワルド・アベリーらの『DNAが遺伝物質であることの実験的証明』を収めた論文が掲載される(J. Exp. Med. 79:137〜158)。この論文はDNA=遺伝物質であることが確実な今、矛盾のないものだが、当時は評価を全く受けなかった(註:この見方は、アヴェリーが属していたロックフェラー研究所およびその周辺での、当初の反響を伝えているに過ぎない。実際には、ジョシュア・レーダーバーグ、ジェームス・ワトソン、マックファーレン・バーネットなど現代遺伝学・分子生物学の元を築いた科学者たちが、「まだ初学者であった頃にアヴェリーらの論文を読んで大きな刺激を受けた」と述べている。ちなみに、ワトソンは彼の著書(ワトソン&ベリー『DNA』)のなかでも、「アヴェリーの実験はハーシーとチェイスの実験が行われる前に既に評価されていた」と重ねて記している。つまり、先を見据えていた科学者の間では正当に評価されていたということである。)
- 1950年 エルヴィン・シャルガフがペーパークロマトグラフィーを用いて塩基存在比に数学的関連があることを明らかにした(AとT、GとCはそれぞれ数が等しいことを示した)。
- 1952年 アルフレッド・ハーシーとマーサ・チェイスによる『ハーシーとチェイスの実験』結果が論文に掲載される(J. Gen. Physiol. 36:39〜56)。本論文によってファージの遺伝物質がDNAであることが確実視されたと言われる。同年、ロザリンド・フランクリンがDNAが二重らせん構造であることを証明するX線回折像写真を撮影する。
- 1953年 ジェームズ・ワトソンとフランシス・クリックによってDNAのB型二重らせん構造のモデルが示され、DNAは生体内で『二重らせん構造』をとっていることを示す論文が発表される(Nature 171:737,738)。
二重らせん構造発見以降の歴史
- 1955年 セベロ・オチョアによってポリヌクレオチドホスホリラーゼが発見された(一見遺伝子とは無関係だが遺伝暗号の解明に寄与した重要な酵素である)
- 1956年 エリオット・ヴォルキンとラザルス・アストラチャンによってDNAからタンパク質への情報のメッセンジャーがRNAである証拠が提出された(mRNAが存在する可能性を示した、このことが確実になったのは5年後、ソル・シュピーゲルマンとベンジャミン・D・ホールらの実験による)
- 1958年 クリックによってセントラルドグマが提唱された(Symp. Soc. Exp. Biol. 12:138〜163)。
- 1959年 ロバート・ホリーによってtRNAala分子が単離された
- 1961年 マーシャル・ニーレンバーグとハインリッヒ・マテイによって大腸菌無細胞発現系を用いたポリウラシルからポリフェニルアラニンが合成される実験が行われた(遺伝暗号解明への初めての実験)。
- 1964年 ニーレンバーグとフィリップ・レーダーによって遺伝暗号解明に大きな寄与をした『トリプレット結合能測定法』が開発された。ヤノフスキーによって遺伝子がタンパク質をコードしていることが示された(遺伝子タンパク質間の共直線性が示された)
- 1966年 遺伝暗号の解読が完了した
- 1970年 ハワード・テミンとデビット・バルチモアがそれぞれある種のウイルスで逆転写反応を見出した(セントラルドグマ概念の訂正)
- 1977年 遺伝子が介在配列によっていくつかの単位に分断されていることが発見された(不連続遺伝子、イントロンの発見)
- 1979年 フレデリック・サンガーによってミトコンドリアではことなる遺伝暗号が使用されていることが発見された(非標準コドンの発見)
- 1981年 トーマス・チェックによって自己スプライシングイントロンが発見された(リボザイムの発見)
脚注
参考文献
- 『細胞の分子生物学』ニュートンプレス - 詳しく知りたい人向け。
- 『Lewin 遺伝子』東京化学同人 - 詳しく知りたい人向け。
関連項目
- 致死遺伝子
- 複対立遺伝子
- 補足遺伝子
- 抑制遺伝子
- 遺伝子組み換え作物
- 遺伝子組換え生物等の使用等の規制による生物の多様性の確保に関する法律(カルタヘナ法)
- 遺伝子診断
- 遺伝子治療
- 遺伝子決定論
- ヒトゲノム計画
- ハプロタイプ
- ナンセンス変異依存mRNA分解機構
- その他は「遺伝子」で始まるページの一覧を参照。
外部リンク
- 遺伝子の部屋(いでんこのへや) - いでんこさんと博士の対話形式で進められる遺伝子についての分りやすい解説。NS遺伝子研究室内。